163
СТАТИЧЕСКОЕ И ДИНАМИЧЕСКОЕ В ЗРИТЕЛЬНОМ
МИРЕ МЛАДЕНЦА
Е. А. СЕРГИЕНКО
Одной из
значительных проблем перцептивной организации человека и ее становления
является проблема отражения статических и динамических характеристик окружающей
среды.
Долгое
время акцент в исследовании этого вопроса ставился на изучении статических
объектов. При таком подходе в моделях и теориях восприятия центральное место
занимал статический мир. Именно поэтому в книге П. Линдсея и Д. Нормана [5]
утверждается, что одна из последних основных задач, которую приходится решать
зрительной системе, — это выделение и оценка движения. Тем не менее накоплено
огромное количество фактов, говорящих о том, что выделение движения — это не
последняя, а генетически первая задача зрительной системы.
В
книге Ш. Ульмана [8] предлагается следующая гипотеза построения системы
представлений об объекте, принципиально отличная от традиционной. Вначале
разные изображения объекта отождествляются друг с другом на основе установления
соответствия при движении, а затем уже выделяется структура представления как
основа понимания, чем же является объект. В традиционной схеме исходными
берутся представления о неподвижном объекте в его различных ракурсах, а затем
возникающая структура используется для анализа движущихся объектов. Однако, как
подчеркивает сам автор, эта гипотеза опирается лишь на теоретические посылки.
Подтверждением могут служить только опытные данные, полученные на генетическом
материале.
В
последние 10—15 лет в области психологии восприятия младенца был накоплен
большой экспериментальный материал, позволяющий пересмотреть представления об
организации его зрительного мира.
Работы
Р. Фантца (Fantz) [9], [12], [13] были
переломными в генетических исследованиях. Его метод зрительного предпочтения
заставил младенца «заговорить» и вызвал новый интерес к онтогенетическому
материалу. Было показано, что младенцы изначально способны к различию
физических характеристик стационарных объектов, перцептивный мир их изначально
организован.
Изучением
восприятия движения у младенцев занимались очень мало (Бауэр [2], Фокс (Fox) [14], Гартлеп (Hartlep) и
некоторые другие). Но и эти немногочисленные исследования не включают движение
в структуру перцептивного мира младенца.
Правомерность
тезиса о примате динамического в зрительном восприятии подтверждается как
фило-, так и онтогенетическим материалом.
Для
животных движение является одним из основных параметров среды. Именно движение
сигнализирует животному об ожидаемой пище или возможной опасности.
Так,
у низших животных величина поля зрения при обнаружении движения становится
определяющим фактором способности выживания. Известно, что амфибии очень плохо
различают неподвижную жертву. Рыбы проявляют интерес только к движущимся
объектам [11]. Шмид, исследуя зрительные реакции собак, показал, что узнавание
подвижных объектов возможно на гораздо больших расстояниях, чем узнавание
неподвижных объектов (цит. по [11]).
Таким
образом, эволюция зрительного восприятия в филогенезе начинается с отражения
динамических свойств, которые потом дополняются отражением других характеристик
окружающей среды.
Само
развитие зрительной системы в онтогенезе предполагает первичность отражения
динамических характеристик.
Младенец
рождается на свет с достаточно полно развитой системой периферического зрения,
доминирующая функция которого заключается в обнаружении движущегося объекта, и
неразвитой центральной областью, отвечающей за детальное восприятие.
Такое
развитие зрительной системы обеспечивает решение жизненно необходимой задачи:
возможности восприятия движения и визуальной дифференцировки изменений во
внешней среде.
Все
изложенные выше соображения привели к постановке экспериментального
исследования, которое должно было выявить последовательность становления
зрительного отражения статики и динамики в раннем онтогенезе человека.
164
МЕТОДИКА
В
эксперименте применялась установка, обеспечивающая движение черно-белых полос
размером 6°. На фоне полос, движущихся со скоростями 28, 20 и 9°/с,
предъявлялись неподвижные стимулы. В качестве неподвижного объекта применялись
два стимула равной величины: в одном случае — схематическое лицо (значимый
стимул), в другом — круг красного цвета (индифферентный, но предпочитаемый
младенцами объект). Размеры объекта составляли 12°, т. е. были равны ширине черной
и белой полос. Момент появления неподвижного стимула отмечался автоматически на
одном из каналов самописца при помощи электромеханического устройства.
Процедура
эксперимента выглядела следующим образом. Младенец лежал в кроватке на спине.
Над ним происходило перемещение полос в течение 10—20 с, затем на фоне движения
предъявлялся один из неподвижных стимулов.
В
экспериментах участвовало 16 детей в возрасте от 4 до 24 недель.
Регистрировались
горизонтальные движения глаз, использовались предусилители УПБ-01 и УБП-02,
самописец Н-327. Скорость движения бумаги самописца составляла 25 мм/с.
РЕЗУЛЬТАТЫ ЭКСПЕРИМЕНТА
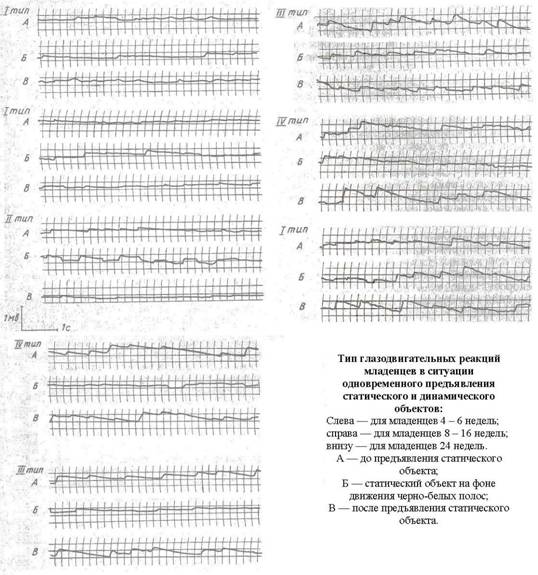
В
условиях описанной задачи были получены следующие типы глазодвигательных
реакций младенцев. Первый тип — все случаи усиления нистагменной активности при
действии статического стимула. В первом случае был зарегистрирован переход
непериодических колебательных движений глаз к появлению выраженной формы
нистагменной реакции. Непериодические колебательные движения глаз в ответ на
перемещение полос наблюдались нами у самых маленьких испытуемых (3—4 недель) и
были классифицированы как первая стадия развития оптокинетического нистагма
(ОКН) в онтогенезе (при исследовании ОКН у младенцев [7]). Во втором случае
появление неподвижного стимула в поле зрения младенца вызывало ОКН даже тогда,
когда ребенок проявлял двигательное беспокойство или кричал,— введение стимула
играло регулятивную роль: успокаивало, направляло его зрительное внимание на
данную задачу, что приводило к глазодвигательной работе в режиме нистагма. И в
третьем случае к первому типу реакции были отнесены собственно оптокинетические
ответы усиления нистагма. Критерием усиления нистагма служило увеличение
частоты и амплитуды глазодвигательных реакций.
Второй
тип реакций на стабильный стимул выражался в разрушении оптокинетической
активности: во время действия стимула наблюдалась замена нистагменных реакций
скачками с последующей фиксацией.
Третий
тип ответов выражался в частичном подавлении нистагменных ответов действием
статического стимула. Основным критерием подавления нистагма являлись изменения
ритма и величины амплитуды.
Четвертый
тип реакций характеризовался полным исчезновением оптокинетических глазных
движений. В периоды предъявления статического стимула регистрировались
фиксационные движения глаз на объекты, иногда сопровождающиеся скачками или
несколькими «ударами» нистагма.
Возрастное
распределение выделенных типов реакций при введении неподвижного стимула на
фоне движущихся полос представлено в табл. 1.
Таблица 1
Типы
глазодвигательных реакций младенцев 4—24 недель на предъявление статических
стимулов на фоне оптокинетических полос
|
Возраст (недели) |
Нистагм |
Процент случаев |
|
4—6 |
I: усиление нистагма II: разрушение нистагма |
60 40 |
|
8—16 |
III: частичное подавление нистагма IV: полное подавление нистагма I: усиление нистагма |
33 36 31 |
|
24 |
III: частичное подавление нистагма IV: полное подавление нистагма |
25 75 |
Из
данных табл. 1 становится ясна динамика изменений реакций младенцев разных
возрастов в данной зрительной задаче. В самом раннем возрасте (4—6 недель, I группа) неподвижный стимул оказывает неспецифическое действие
на зрительную
165
систему
младенцев, в то время как у взрослых предъявление статического объекта на фоне
движущихся полос вызывает либо полное, либо частичное подавление ОКН.
Доминирующей реакцией в такой ситуации у детей самой младшей группы является
усиление ОКН.
У
младенцев 8—16 недель (II группа) основной
удельный вес приобретает уже специфическая «взрослая» реакция — подавление ОКН,
которая отсутствует у совсем маленьких детей, тогда как число неспецифических
реакций — усиления ОКН при неподвижном стимуле — уменьшается в два раза.
В III возрастной группе (24 недели) несомненна однотипность
реакций на статический стимул, предъявляемый на фоне движущихся полос это подавление
нистагменных ответов. По степени этого подавления можно выделить два типа
реакций. Третий тип — предъявление статического стимула частично подавляет
оптокинетическую активность. К этому типу относятся случаи возникновения саккад
при предъявлении стимула вправо и влево с последующей фиксацией.
В III группе младенцев реакций усиления ОКН действием
статической стимуляции не обнаружено. Реакция подавления у детей 24 недель
составляет 100 % всех ответов, тогда как у детей 8—16 недель — 69 %. Особенно сильно
увеличивается число реакций полного подавления нистагменной активности (четвертый
тип реакций):
166
во II группе они составляли 36%, а в III — 75 %, т.е. в два раза чаще встречались у детей 24 недель. На
рис. 1 представлены типичные записи движения глаз у младенцев 4—24 недель в
данной зрительной задаче.
В
принципе по действию статического стимула на зрительную систему младенцев все
ответы детей исследуемого возраста можно свести к двум основным реакциям,
определяющим избирательность младенцев: специфическая реакция, когда
статический объект доминирует в восприятии, и неспецифическая — движение полос
оказывается наиболее значимой стимуляцией.
Выделенные
типы ответов являются не только возрастной характеристикой. Все реакции,
свойственные данному возрасту, могут встречаться у одного и того же
испытуемого, в пределах одной экспериментальной программы. На появление того
или иного типа реакций влияют такие факторы, как скорость движения полотна,
число предъявлений статического стимула (т.е. продолжительность эксперимента),
характер предъявляемого объекта. В табл. 2 показана зависимость появления
разного типа ответов от скорости перемещения полос для младенцев 8—16 недель.
При
быстрых скоростях движения полотна реакции «взрослого» типа значительно
преобладают над ответами при медленной скорости движения (сравните: 82 %
подавления ОКН полное и частичное при скорости движения полос 28°/с и 60 % —
при 9°/с). В то же время удельный вес неспецифических ответов, т.е. усиление
ОКН при переходе к медленной скорости движения полос возрастает в 2,2 раза.
Скорость движения полос не оказывает такого влияния на характер зрительных
реакций у младенцев как младшей (4—6 недель), так и старшей группы (24 недели).
Таблица 2
Количество
реакций разных типов при различных скоростях движения полотна у детей 8—16
недель
|
Скорость движения полос (7с) |
Тип реакции |
|||
|
|
Частичное подавление ОКН |
Полное подавление ОКН |
Подавление ОКН (суммарно-полное и частичное) |
Усиление ОКН |
|
28 20 9 |
44 21 40 |
38 42 20 |
82 63 60 |
18 37 40 |
Наблюдается
некоторое изменение соотношения различных типов ответов в порядке предъявления
стимула. Эти изменения носят волнообразный характер и зависят, по-видимому, от
колебаний внимания к данной зрительной задаче.
Самое
сильное влияние на появление того или иного типа реакций оказывает характер
предъявляемого объекта. В табл. 3 дано количество выделенных типов реакций при
действии значимого («лицо») и индифферентного стимула (красный круг).
Таблица 3
Число разных
типов реакций на значимый и индифферентный стимул, предъявляемый на фоне
движущихся полос у младенцев 4—24 недель (%)
|
Возраст (недели) |
Тип реакции |
Значимый стимул («лицо») |
Индифферентный стимул (красный круг) |
|
4—6 |
I: усиление ОКН II: «разрушение» ОКН |
18 37 |
27 18 |
|
8—16 |
III: частичное подавление ОКН IV: полное подавление ОКН I: усиление ОКН |
14 38 3 |
17 16 14 |
|
24 |
III: частичное подавление ОКН IV: полное подавление ОКН |
5 50 |
20 25 |
|
4—6 8—16 24 |
По всем типам По всем типам По всем типам |
55 55 55 |
45 45 45 |
Самые
существенные различия в зависимости от характера стимуляции наблюдаются при
появлении реакций первого типа (усиление нистагменной активности). Если
предъявление значимого стимула вызывало ответы первого типа в 3 % случаев, то
появление индифферентного — в 14 % случаев у детей 8—16-недельного возраста.
Таким образом, действие значимого стимула в 4,6 раза менее эффективно для
«раскачки» нистагма, чем действие индифферентного, в то же время значимый
стимул вызывает полное подавление нистагменной активности в 2,5 раза чаще, чем
индифферентный стимул. Особенно следует подчеркнуть этот факт в связи с тем, что
именно в
167
возрасте 8—16
недель наиболее интенсивно развивается комплекс оживления у младенцев, и
поэтому схематическое изображение лица человека, появляющееся в поле зрения, в
значительной мере опосредует зрительные реакции детей этого возраста.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Данная
экспериментальная задача ставит младенца перед выбором зрительной стимуляции —
фиксировать статический объект или реагировать на движение оптокинетической
стимуляции. Фиксационный рефлекс и рефлекс на движение объектов как
физиологические предпосылки отражения данной визуальной задачи являются
врожденными зрительными реакциями, однако рефлекс на движение объекта —
филогенетически более древний. В то же время оптокинетические реакции у
младенцев вызвать легче, чем прослеживание движущегося объекта или простую
фиксацию его [15]. Оптокинетический нистагм можно обнаружить у всех животных,
имеющих подвижное глазное яблоко [19].
Все
эти факты приводят к заключению, что оптокинетическая стимуляция (движение
контрастов) является филогенетически более древним видом движущейся стимуляции,
выделяемой зрительной системой.
Экспериментальную
ситуацию, использованную в наших исследованиях, можно представить как модель
развития зрительной системы человека: филогенетически более древние функции
зрения — отражение движения контрастов — вступают «в соревнование» с более
новыми функциями — отражение статического объекта. Основные итоги исследования
данной модели в раннем онтогенезе показали:
1.
Движущиеся стимулы на самых ранних этапах младенчества являются наиболее
значимыми для ребенка.
Статический
объект у детей 4—6 недель вызывал лишь усиление реакции на движение, т.е.
оказывал неспецифическое действие на зрительную систему — активизировал внимание
к предпочитаемой подвижной стимуляции.
В
возрасте 8—24 недель статический стимул начинает оказывать адекватное действие.
Он подавляет реакции на движение, причем биологически значимый объект («лицо») вызывает
полное переключение зрительной системы на статический стимул в 2,5 раза чаще,
чем индифферентный.
2. Мы
регистрировали два типа ответов: а) ОКН, б) фиксационные движения глаз на
неподвижный объект. На всем исследуемом отрезке онтогенетического развития обе
эти реакции претерпевают изменения, связанные с созреванием зрительной системы
и переходом на новый уровень зрительной обработки.
Выделение
признаков сенсорного сигнала происходит путем многоканальной, многоуровневой
переработки информации. Чем выше уровень, тем больше количество выделяемых
признаков, тем точнее их анализ. Обработка жизненно важных сигналов, требующая
скорейшего двигательного ответа, осуществляется за счет работы более низких
уровней организации нервной системы. Поэтому на низком уровне организации
сенсомоторной интеграции в первую очередь становится возможным отражение самых
грубых общих, жизненно необходимых параметров окружения: движения,
контрастности, значимости. Процесс постепенного совершенствования зрительных
функций приводит к формированию более высоких уровней отражения, что выражается
в более полном и точном описании окружения.
Внутренней
предпосылкой перехода от низких уровней организации зрения к более высоким в
онтогенезе является созревание. Несомненна гетерохрония в развитии зрительной
системы как в морфологическом, так и функциональном отношении.
К
моменту рождения ребенка оказываются сформированными филогенетически более
древние структуры зрительной системы: хорошо развита периферия сетчатки,
фовеальная область находится в стадии созревания, центральная регуляция
обеспечивается низким уровнем — стволо-ретикулярным [3]. К четырехмесячному
возрасту заканчивается дифференцировка структур сетчатки и продолжается
активное развитие центрального звена зрительной системы, охватывающее все более
высокие «этажи» мозговой организации. Прижизненное развитие наиболее сложных
функций зрительной системы имеет принципиальное значение для человека: этим
достигается одно из самых важных ее качеств — пластичность в работе.
Гетерохрония
в развитии зрительной системы и определяет построение как сенсорных, так и
моторных компонентов зрения на разных этапах онтогенеза. Доминирование в
становлении системы зрения функции периферической субсистемы обуславливает
возможности отражения на самых ранних этапах развития младенца. Именно поэтому
дети 4—6 недель предпочитают движущиеся полосы статическому объекту.
Неподвижный
стимул, действуя на развивающуюся фовеальную субсистему, приводит в этот период
лишь к дополнительной
168
адеквации
доминирующей субсистемы периферического зрения. Этим объясняется усиление
нистагменных реакций у самых маленьких детей (4—6 недель) при предъявлении
статических объектов.
Выраженная
реакция на статический стимул как раз приходится на период активного созревания
центральной зрительной субсистемы. Дополнительным свидетельством перестройки
зрительного аппарата служит изменение параметров ОКН. Если в 4—6 недель ОКН
строится на самом низком уровне, о чем свидетельствуют его характеристики:
малоамплитудная, тоническая реакция — скорость прослеживающих движений глаз не
зависит от скорости движения полос (подробный анализ характеристик ОКН у
младенцев дан в работе автора [7]). ОКН такого типа может быть охарактеризован
как «несмотрящий», периферический нистагм. Подобный ОКН можно обнаружить у всех
животных, имеющих подвижное глазное яблоко. Что это означает? Что на самых
ранних стадиях развития зрительная система обеспечивает самую общую, грубую,
приблизительную обработку визуальной информации, в данном случае динамической
ситуации.
Следующий
этап развития (8—24 недель) характеризуется становлением «смотрящего»,
центрального нистагма (с выраженной амплитудой, четким ритмом, появлением
соответствия между скоростью прослеживающей фазы и движением полос). Такая
форма нистагма требует участия центральной зрительной субсистемы, которая
обеспечивает детальную обработку сигнала. Таким образом происходит построение
уровня точной обработки информации.
Особого
объяснения требует факт, обнаруженный нами у детей 8—16 недель (на стадии
активного формирования фовеального зрения). В этой группе все еще встречаются
неспецифические реакции на статический объект, предъявленный на фоне движущихся
полос, причем при снижении скорости перемещения оптокинетических стимулов число
таких «инфантильных» реакций значительно возрастает. Здесь необходимо
напомнить, что фовеальная и периферическая субсистемы находятся в реципрокных
отношениях. Предположительно эти отношения начинают складываться как раз в
период 8—16 недель, когда центральная субсистема достигает такого уровня
развития, который уже может конкурировать с хорошо развитой периферической
субсистемой. В возрасте 4—6 недель неразвитое центральное зрение не может
подавить периферические функции — отсюда возникают неспецифические ответы зрительной
системы. В то же время известно, что периферия сетчатки по своей организации
неоднородна как морфологически, так и функционально. Чем эксцентричнее
расположены рецептивные поля, тем больше их размеры и соответственно выше
должна быть скорость движения стимулов для их активации [10], [17]. По нашей
гипотезе, развивающиеся реципрокные отношения центральной и периферической
областей сетчатки более выражены тогда, когда сильнее активизированы дальние
области периферии с большими рецептивными полями, реагирующими на высокие
скорости движения. При достаточном удалении «очагов» активации различия в специфике
функций центральной и периферической зон становятся более выраженными. При
медленных же скоростях движения стимула задействованы ближайшие к фовеа области
периферии сетчатки, и дифференцировка в активации субсистем затруднена, что и
приводит к появлению неспецифических «инфантильных» реакций усиления ОКН. Такие
реакции отсутствуют у младенцев 24-недельного возраста, когда зрительная
система морфологически и функционально становится достаточно зрелой, с
определенными реципрокными отношениями субсистем.
В
целом раннему периоду онтогенетического развития свойственно наличие грубых
дифференцировок; по нашим представлениям, эта закономерность также проявляется
и в развитии зрительной системы.
Переход
от уровня самых грубых и общих оценок к уровню точного анализа осуществляется в
силу гетерохронного развития зрительной системы.
Процесс
зрительного восприятия можно представить как процесс уровневой фильтрации,
который привносит свои особенности в специфику отражения внешних параметров
данного уровня.
Данная
концептуальная схема подтверждается многими нейрофизиологическими данными. Так,
например, в работе Ф.А. Ата-Мурадовой [1] показана иерархия развития мозговых
структур, где каждый уровень структурно и функционально обеспечивает отражение
тех характеристик, которые жизненно необходимы на данном уровне развития
организма.
Какими
же механизмами обеспечивается работа самого низшего уровня? Первичный анализ
зрительных задач осуществляется, по всей вероятности, генетически закрепленными
механизмами.
В
задачу настоящего сообщения не входило доказательство этого тезиса, однако
укажем, что экспериментальные данные, полученные сотрудниками группы по
изучению
169
онтогенеза
перцептивных процессов и другими авторами, свидетельствуют в пользу этого
положения.
Приведем
лишь некоторые факты:
1) врожденность
механизма саккад и плавной формы ГДА, т.е. основных форм моторики глаза,
обеспечивающие все сенсомоторные акты зрения [4],
2) врожденность
зрительно-вестибулярных [6], зрительно-слуховых взаимодействий, обеспечивающих
общую сенсомоторную оценку пространства.
Резюмируя
все сказанное, необходимо подчеркнуть: особенности эволюционного развития
функций зрительной системы от выделения движения до анализа и статических
объектов сохранились в последовательности развития функций зрительной системы
человека.
1. Ата-Мурадова
Ф. А. Развивающийся мозг: системный анализ. Генетические детерминанты. М.,
1980. 295 с.
2. Бауэр
Т. Психическое развитие младенца. М., 1979. С. 217—292.
3.
Возрастная физиология // В серии: Руководство по физиологии. М., 1975. 328 с.
4.
Движение глаз и зрительное восприятие. М., 1978. С. 9—71.
5. Линдсей
П., Норман Д. Переработка информации у человека. М., 1974. С. 159—175.
6. Митькин
А. А. Зрительно-вестибулярное взаимодействие у младенцев // Психол. журн.
1981. Т. 2. № 4. С. 68—80.
7. Сергиенко
Е.А. Ранний онтогенез глазодвигательной активности в условиях динамической
стимуляции: Автореф. канд. дис. М., 1978. 24 с.
8. Ульман
Ш. Принципы восприятия подвижных объектов. М., 1983. 169 с.
9. Фанц
Р. Восприятие формы // Восприятие. Механизмы и модели. М., 1974. С.
338—351.
10. Ярбус
A. Л., Рожкова
Г. И. Особенности
восприятия объектов на периферии поля зрения // Сенсорные системы. М.,
1977. С.
64—73.
11. Duke-Elder S. System of ophthalmology. V.
1. The eye in evolution. Sant Luis, 1958.
12. Fantz R. Configurational selectivity: Critical
for development of visual perception and attention // J. Psychol. 1979. V. 33
(4). P. 277— 287.
13. Fantz R. Pattern discrimination and
selective attention as determinants of perceptual development in children. N.Y.,
1966.
14. Fox R., McDaniel С. The
perception of biological motion by human infant // Science. 1982. V. 218. P.
486—487.
15. German G. G., Cogan D. A., Gellis S. S. An
apparatus for grading the basic of optokinetic nystagmus // Pediatrics. 1957.
V. 19. P. 1088— 1092.
Поступила в
редакцию 14.VI 1984 г.