95
ИНДИВИДУАЛЬНЫЕ РАЗЛИЧИЯ В РЕАКЦИЯХ НА ИНДИФФЕРЕНТНЫЕ И НЕПРИЯТНЫЕ СТИМУЛЫ
Э. М. РУТМАН, Б. И. КОЧУБЕЙ, Ю. Н. ШЕВАРЕВ
После исследований Е. Н. Соколова [8], выделившего два типа реакций на новый или неожиданный стимул — ориентировочные (ОР) и оборонительные (ОбР), было проведено огромное количество исследований ОР в самых разных контекстах. Детально исследовались изменения разных показателей в ходе ОР и их взаимосвязи, механизм возникновения и угашения ОР, индивидуальные различия ОР и их связь с психологическими особенностями. Было показано, в частности, что индивидуальные особенности ОР коррелируют с особенностями выработки простых и сложных условных рефлексов [7], [8], [29] с показателями обучения [2], [28] и деятельности [20], с психологическими характеристиками личности [20], [24]. ОР стали рассматривать как наиболее простой случай организации нового действия [6], а ее угашение — как элементарную модель обучения [8], [11]. В характеристиках ОР стали искать индикаторы индивидуальных особенностей, относительно независимые от языка и культуры. Например, была обнаружена корреляция между скоростью снижения частоты пульса при повторении нового зрительного стимула у детей в возрасте 9 мес. с показателями их интеллекта в четырехлетнем возрасте [29]. Эти данные позволяют надеяться, что ОР может служить инструментом исследования гено- и паратипических факторов формирования индивидуальных особенностей, коррелирующих с характеристиками ОР.
ОбР исследовалась несравненно меньше. Условную ОбР нередко используют при поисках физиологических индикаторов индивидуальных особенностей «стиля преодоления» трудной ситуации [13], [14]. При изучении влияния когнитивных факторов на течение стресса ОбР успешно используется в качестве простейшей модели реакции на стресс [14]. По-видимому, ОбР, аналогично ОР, может служить инструментом исследования соотношения средовых и
96
наследственных факторов в прижизненном формировании индивидуальных особенностей поведения в стрессогенных условиях. Для этого нам представляется более целесообразным изучать характеристики ОбР на безусловный, а не на условный стимул, поскольку применение последнего вносит дополнительную переменную — индивидуальные особенности выработки условного рефлекса. В качестве безусловного стимула чаще всего применяется электрокожная стимуляция, реже — громкий звук. Применение громкого звука кажется более адекватным, поскольку не осложняется широко распространенным страхом перед ударом тока.
Характеристики ОбР на громкий звук изучены гораздо меньше, чем ОР на звуки средней интенсивности. В качестве признаков, отличающих ОбР от ОР, указываются увеличение частоты пульса [33] и частоты межстимульных миганий [30]. Еще меньше изучена динамика привыкания при ОбР1.
Преобладает общее представление, что привыкание при действии «оборонительных» стимулов по сравнению с ОР замедляется, уменьшается или вовсе не возникает [22]. Уменьшение или прекращение привыкания при повышении интенсивности стимула рассматривается как один из основных критериев привыкания в работе Томпсона и Спенсера [37]. Однако есть данные об увеличении и ускорении привыкания при повышении интенсивности стимуляции [21], [39]. Детальный обзор исследований связи привыкания с интенсивностью стимуляции [21] показал, что связь привыкания с интенсивностью неоднозначна для разных показателей и не имеет характера простой отрицательной связи, как предполагали Томпсон и Спенсер. Более того, автор даже пришел к выводу о прямой связи привыкания с интенсивностью при ОбР. Таким образом, не существует единого мнения о закономерностях привыкания при ОбР и отличии их от привыкания при ОР.
Исходя из этого, мы предприняли исследование изменений пульса, КГР, миганий и вызванных потенциалов (ВП) при повторении громкого звука, вызывающего ОбР, и звука средней интенсивности, вызывающего ОР. Кроме самих названных показателей мы исследовали их взаимосвязи при ОбР и ОР, а также взаимосвязь величины и длительности привыкания с величиной начальной реакции по каждому из показателей и между разными показателями.
МЕТОДИКА
Данная работа является частью более широкого исследования соотношения наследственных и средовых факторов в характеристиках ОР и ОбР близнецовым методом, поэтому в экспериментах принимали участие 40 здоровых близнецов (9 моно- и 11 дизиготных пар) в возрасте 19—29 лет. Во время опыта испытуемые сидели с закрытыми глазами в удобном кресле в темной звукоизолированной комнате. Предварительно им говорили, что исследуется их биоэлектрическая активность, от них не требуется никаких реакций на возможные в связи с проверкой приборов сигналы. Вначале предъявляли с интервалом в 10 с 33 раза тон частотой 1000 Гц, громкостью 80 дБ над уровнем 0,0002 бара, длительностью 480 мс. Затем, через 30 с, с тем же межстимульным интервалом предъявляли 33 раза тон той же частоты и длительности громкостью 105 дБ. Регистрировали ЭЭГ в точке Cz ЭОГ, КГР и ЭКГ. Регистрация ЭЭГ велась монополярно с объединенным ушным электродом на электроэнцефалографе Nihon Koden, постоянная времени 0,1, фильтр высоких частот — 25 Гц. Усреднение ВП производилось на ЭВМ «АТАС-350», период анализа—1024 с, начало усреднения — за 256 мс до стимула. Усредняли отдельно ВП на 1 — 10-й, 4—13-й, 11—20-й и 21—30-й стимулы, параллельно производилось усреднение ЭОГ для последующей корреляции возможных артефактов, связанных с миганием, по разработанной ранее методике [20].
АНАЛИЗИРУЕМЫЕ ПОКАЗАТЕЛИ
Показатели ВП. 1. Амплитуда ВП 1 — 10ОР— амплитуда от пика О105 до пика П190 в мкв ВП, усредненного по ответам на 1 — 10-й звуки 80 дБ.
2. Амплитуда ВП 1—10ОбР — тот же показатель для звука 105 дБ.
3. Прирост амплитуды ВП ОбР — разность между показателями 2 и 1.
4. Привыкание ВП ОР. В качестве показателя привыкания ВП вычислялся коэффициент в линейной зависимости у = a + bх [9], где х — среднее арифметическое взятых для усреднения номеров ответов на стимулы 80 дБ (например, при усреднении ответов с 1-го по 10-й х = 5,5), у — нормированное значение амплитуды О1 — П2, где компонент О1 имел латентность 105,1 ± 2,89 мс, а П2— 184+4,56 мс.
5. Привыкание ВП Об —тот же показатель для ВП на звук 105 дБ.
6. Прирост привыкания ВП Об / ОР — разность между показателями 5 и 4.
ПОКАЗАТЕЛИ ЭКГ
7. Изменения ЧСС 1—10 ОР — изменение среднего значения интервала между зубцами R ЭКГ в мс в период предъявления 1—10-го звуков 80 дБ по сравнению с тем же показателем в фоне (до стимуляции). Положительная величина означает укорочение интервала.
8. Изменения ЧСС 1—10 Об — тот же показатель для звука 105 дБ.
97
9. Прирост изменений ЧСС Об/ ОР — разность между показателями 7 и 8.
10. Привыкание ЧССОР—разность между средней ЧСС в период предъявления 21—30 звуков 80 дБ и средней ЧСС при предъявлении 1—10 звуков той же интенсивности.
11. Привыкание ЧСС Об — тот же показатель для звуков 105 дБ.
ПОКАЗАТЕЛИ КГР
12. Амплитуда КГР 1ОР — амплитуда, измеряемая в мкв от максимума отрицательного до максимума положительного колебания КГР на 1-й звук 80 дБ.
13. Амплитуда КГР 1Об — тот же показатель для звука 105 дБ.
14. Прирост амплитуды КГР Об/ОР — разность между показателями 12 и 13.
15. Привыкание КГР ОР — амплитуда КГР на 10-й звук 80 дБ в % от величины амплитуды КГР на 1-й звук 80 дБ.
16. Привыкание КГРОб — тот же показатель для звука 105 дБ.
17. Длительность привыкания КГР ОР — количество звуков 80 дБ до отсутствия КГР на три стимула подряд.
18. Длительность привыкания КГР Об — то же для звуков 105 дБ.
19. Прирост длительности привыкания КГРОб — разность между показателями 17 и 18.
ПОКАЗАТЕЛИ МИГАНИЯ
20. Учащение МСМ 1—10 ОР — величина прироста среднего значения частоты МСМ в период предъявления 1—10-го звуков 80 дБ по сравнению с величиной мигания в фоне (до стимуляции).
21. Учащение МСМ 1—10 Об —тот же показатель для звуков 105 дБ.
22. Прирост учащения МСМ Об — разность между показателями 20 и 21.
23. Привыкание МСМ ОР — разность между величиной учащения МСМ в период предъявления 20—30-го звуков 80 дБ по сравнению с фоном и тем же показателем для 1—10-го звуков 80 дБ.
24. Привыкание МСМ Об — показатель, аналогичный предыдущему, для звука 105 дБ.
25. Длительность привыкания МР — количество стимулов до угашения мигательного рефлекса в ответ на звук 105 дБ.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
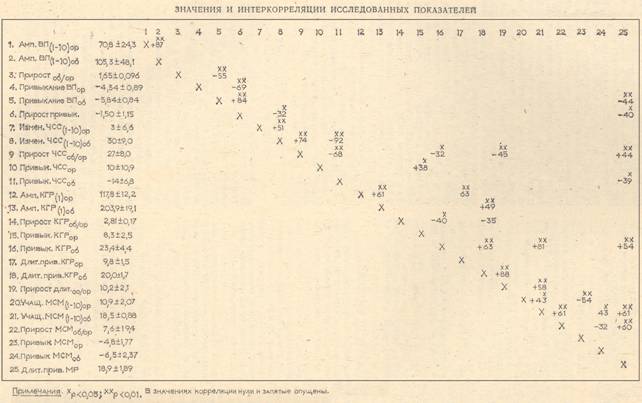
Характеристики ОР и ОбР. Средние значения и стандартные отклонения всех регистрируемых показателей представлены в таблице. Статистически достоверными при переходе от звука 80 дБ к звуку 105 дБ оказались следующие изменения: увеличение амплитуды КГР (t = 5,11, p<0,01), увеличение числа стимулов до угашения КГР (t = 5,13, p<0,001), увеличение амплитуды ВП (t = 6,7, р<0,001), ЧСС (t=3,63, p<0,001), частоты МСМ (t=3,85, р<0,01) и привыкания МСМ (t = 2,05, p<0,05>. Звук 105 дБ вызывал мигательный рефлекс, исчезавший, в среднем, после 19 стимулов. Увеличение ЧСС, учащение МСМ и появление мигательного рефлекса свидетельствуют о том, что звук 105 дБ в отличие от звука 80 дБ вызывал ОбР, в соответствии с данными ряда исследований [8], [14], [19], [34]. В отличие от этого тон 80 дБ практически не вызывал (в среднем по группе) увеличения ЧСС, не вызывал МР. В соответствии с данными, полученными у ряда авторов [8], [32], [33], мы считали, что этот стимул вызывает ОР.
Наиболее интересным представляется тот факт, что при переходе к ОбР ни по одному показателю, кроме длительности привыкания КГР, не наблюдается уменьшения привыкания по сравнению с ОР. Более того, по ВП и частоте МСМ происходит увеличение привыкания, достигающее достоверности только для МСМ. Таким образом, полученные результаты наряду с другими, упоминавшимися выше данными, не подтверждают распространенное представление об ухудшении привыкания при ОбР и демонстрируют разные изменения привыкания по разным показателям.
При переходе от ОР к ОбР отмечены также статистически достоверные изменения дисперсий: увеличение дисперсии амплитуды КГР на первый стимул (F=2,45) и величины привыкания КГР (F = 3,10), уменьшение дисперсии частоты МСМ (F = 5,53) и привыкания ЧСС (F = 2,57). Отмеченные различия, особенно уменьшение дисперсии частоты МСМ при увеличении средней величины в свете данных о связи частоты МСМ с эмоционально отрицательной активацией [36], подкрепляют представление о разных механизмах активации в случае ОР и ОбР [21], [31]. В связи с отмеченным уменьшением межиндивидуальной дисперсии некоторых показателей при ОбР стоит упомянуть, что в ряде исследований [25], [35] отмечалось увеличение внутрииндивидуальной стабильности показателей при повторных экспериментах в случае звуков высокой интенсивности (например, после 90 дБ по сравнению с 70 дБ).
АНАЛИЗ ПОЛУЧЕННЫХ ИНТЕРКОРРЕЛЯЦИЙ
1. Показатели ВП. Высокая положительная связь амплитуд ВП на звуки 801 и 105 дБ ( + 87, см. табл.) отражает тенденцию к сохранению испытуемыми их ранговых мест при переходе от ОР к ОбР. Чрезвычайно интересной представляется отрицательная связь (—55) между привыканием ВП при ОбР и величиной прироста амплитуды ВП при ОбР по сравнению с ОР. Иначе говоря, у лиц с наибольшим приростом амплитуды ВП при ОбР по сравнению с ОР наблюдается тенденция к более выраженному привыканию ВП.
2. Показатели ЭКГ. Как и в случае ВП, отмечается тенденция к сохранению испытуемыми своих ранговых мест по степени увеличения ЧСС в ответ на звук 80 дБ при переходе к звуку 105 дБ. Между величиной начальной реакции и привыканием ЧСС при ОбР существует такая же связь, какая отмечена выше для ВП. Отрицательные корреляции —92 и —68 (см. табл., ст. 11)2 указывают, что привыкание ЧСС при ОбР больше выражено у тех испытуемых, которые отличались большим увеличением ЧСС после звуков 105 дБ как по абсолютной величине, так и по сравнению с ЧСС после звуков 80 дБ.
98
99
3. Показатели КГР. Положительные корреляции + 61 и +88 отражают тенденцию к сохранению испытуемыми ранговых мест по амплитуде КГР и длительности привыкания в разных ситуациях, как это было отмечено для амплитуды ВП и величины изменения ЧСС. Корреляции +63 и +49 (см. табл., стр. 12 и 13) показывают, что у испытуемых с наибольшей амплитудой КГР отмечается большее число стимулов до полного прекращения реакции как после звука 80 дБ, так и после 105 дБ, что может объясняться морфофункциональными характеристиками периферического (кожного) аппарата генерации КГР (количество потовых желез, их активность и т. д.). Особый интерес представляют отрицательные корреляции —40 и —35 (стр. 14), которые показывают, что самое быстрое и максимальное по величине привыкание отмечается у лиц с наибольшим приростом амплитуды КГР при ОбР по сравнению с ОР. Стоит также отметить, что при ОбР у испытуемых с наибольшим по величине привыканием отмечается тенденция к наиболее быстрому привыканию КГР (корреляция +63, стр. 16), а при ОР подобной взаимосвязи нет. Таким образом, в случае КГР, как и для ранее рассмотренных показателей, увеличение привыкания у лиц с наиболее выраженными начальными реакциями отмечается только в случае ОбР.
4. Показатели мигания. Как и по всем другим показателям, обнаруживается тенденция к сохранению испытуемыми ранговых мест по учащению МСМ после звуков 80 и 105 дБ (+43). Отрицательные корреляции показывают тенденцию к увеличению привыкания при ОбР у лиц с наиболее выраженной начальной реакцией, измеряемой по абсолютной величине (—43) или по изменению в сравнении с ОР (—32). Наблюдается более выраженное привыкание у лиц с наиболее выраженным учащением МСМ также и при ОР (—54). Стоит отметить, что у лиц с наибольшим учащением МСМ при ОбР отмечается тенденция к наибольшей длительности привыкания МР ( + 60 и+ 61, см. табл., стр. 25). Отсутствие такой корреляции для МСМ при ОР позволяет думать о появлении нового фактора при ОбР, влияющего на частоту МСМ и длительность привыкания МР.
5. «Межсистемные» корреляции. Корреляции между разными показателями КГР, ВП, ЧСС и миганий мы условно будем называть межсистемными. Прежде всего представляет интерес само наличие «межсистемных» корреляций, поскольку в известных нам подобных исследованиях при стимулах средней интенсивности таких корреляций не было [7], [25]. При анализе «межсистемных» корреляций вновь отчетливо выявляются различия между ОР и ОбР. Так, все положительные «межсистемные» корреляции отмечаются только при ОР и отражают, по-видимому, уровень активации. Подобных корреляций при ОбР не обнаруживается. Например, при ОР привыкание КГР больше у испытуемых с наименьшим увеличением ЧСС ( + 3S), а при ОбР, наоборот, привыкание КГР больше у испытуемых с наибольшим приростом ЧСС после звука 105 дБ по сравнению с ЧСС после звука 80 дБ (—32, ст. 16). Иначе говоря, при ОР привыкание КГР выражено больше у лиц с наименьшей, а при ОбР — с наибольшей начальной реакцией ЧСС,- что говорит о разной природе снижения величины реакций при повторении стимула в случае ОР и ОбР. При «межсистемном» сопоставлении большая часть корреляций при ОбР обнаруживает ту же закономерность, что и при анализе внутри системы одного и того же показателя: увеличение привыкания при ОбР по одному показателю выражено больше у лиц с наиболее выраженной начальной реакцией по другому показателю. Так, тенденция к более быстрому привыканию КГР отмечается у лиц с наибольшим приростом ЧСС при ОбР по сравнению с ОР (—45, стр. 9). Привыкание и прирост привыкания ВП, привыкание ЧСС при ОбР выражены больше у лиц с наиболее длительным МР (—44, —40 и —39, ст. 25). Прирост привыкания ВП при ОбР выражен больше у лиц с наибольшим увеличением ЧСС после звука 105 дБ (—32, стр.6).
Выявленные взаимосвязи между привыканием в одной «системе» и выраженностью ОбР по показателю другой «системы» снимают возможные методические возражения, возникающие при интерпретации связи привыкания с величиной начальной реакции «внутри» одного и того же показателя (см. подробно [25]). . Это позволяет с уверенностью утверждать, что у испытуемых с более выраженной оборонительной реакцией наблюдается тенденция к увеличению привыкания. Подобная взаимосвязь между привыканием и начальной реакцией при ОР наблюдается только в одном случае — для привыкания МСМ.
Нам кажется, что полученные результаты перекликаются с экспериментальными данными и представлениями, развиваемыми в серии работ Бухсбаума с соавторами [18]. Согласно этим работам, при повышении интенсивности стимула начиная с некоторого уровня амплитуда ВП у части испытуемых перестает увеличиваться или даже снижается. На основании этого факта было выдвинуто предположение о наличии в ц. н. с. защитного, компенсаторного механизма, снижающего величину реакций при действии слишком интенсивных для данного индивидуума стимулов. Этот механизм мыслится как некое подобие «сенсорного фильтра» отдельно для каждой модальности. Мы полагаем, на основании рассмотренных выше данных, а также данных об увеличении степени снижения амплитуды ВП на стимулы высокой интенсивности при повышении уровня активации [16], что существует не «сенсорный фильтр», а модально-неспецифический защитный механизм, который включается в связи с оборонительной реакцией. Высокая интенсивность включает этот механизм лишь постольку, поскольку вызывает оборонительную реакцию. В пользу этого предположения говорит и тот факт, что увеличение привыкания при ОбР связано в большей части случаев именно с приростом величины реакции по сравнению с ОР, а не с абсолютным ее значением. Рассматривая этот прирост как показатель сдвига функционального состояния, связанный с переходом от ОР к ОбР, мы предполагаем, что именно выраженность ОбР определяет степень включения защитного механизма, приводящего к снижению величины реакций. Это подкрепляется и такими данными [1], как изменения порога МР в ответ на звук при изменениях
100
эмоционального состояния при изменениях порога восприятия звука. Разная связь привыкания с интенсивностью для стимулов средней и высокой интенсивности отмечалась в обзоре Грэхем [21]. О различии привыкания при ОР и ОбР говорит тот факт, что ни по одному показателю мы не обнаружили корреляции между привыканием при ОР и ОбР.
Высказанному предположению об увеличении привыкания при увеличении ОбР соответствуют также данные о снижении ВП на стимулы невысокой интенсивности, но содержащие неприятную информацию [3], об уменьшении амплитуды КГР при увеличении интенсивности стимула в случае очень высокого исходного уровня активации [11], а также многочисленные данные, полученные в русле исследований так называемой перцептивной защиты [17].
Предположение о включении при действии оборонительного стимула компенсаторного механизма, направленного на снижение реакций при повторном действии стимула, соответствует существующим представлениям о том, что одной из функций ОбР является снижение чувствительности к неприятным или угрожающим стимулам, если их невозможно избежать [8], [35].
Не исключено, однако, и другое, «когнитивное» объяснение полученных результатов. Хорошо известно, что ожидание неприятного стимула при знании момента его предъявления приводит к снижению вегетативных реакций [32], амплитуды ВП и МР [1] и субъективных оценок степени неприятности или интенсивности стимула [38]. В то же время показано, что чем более неприятны стимулы или чем более тревожны испытуемые, тем больше «включается» ожидание стимула, усиливается поиск информации, позволяющей подготовиться к неприятному стимулу, предвидеть момент его предъявления [10], [14], [23]. Не исключено, что полученные корреляции привыкания с начальной величиной ОбР обусловлены тем, что лица с более выраженной ОбР больше старались предвидеть момент предъявления стимула, подготовиться к нему, что и обеспечивало большее и более быстрое снижение реакций при повторении. О роли «когнитивного» механизма говорит обнаруженный нами ранее [5] факт зависимости связи между привыканием и величиной реакции ВП и МСМ от отношения испытуемых к эксперименту. При «плохом» отношении высокая амплитуда ВП и частота МСМ сочетались не с увеличением, а с уменьшением привыкания. Возможные причинно-следственные механизмы этой связи подробно рассмотрены в упомянутой работе. Обе интерпретации не являются взаимоисключающими.
1. Волынкина Г. Ю., Суворов Н. Ф. Нейрофизиологическая структура эмоциональных состояний человека. — Л., 1981.
2. Воронин Л. Г., Соколов Е. Н., У Баохуа. Типологические особенности ориентировочного рефлекса у человека. — Вопросы психологии, 1959, № 6.
3. Костандов Э. А. Восприятие и эмоции. — М., 1977.
4. Кочубей Б. И., Шеварев Ю. Н. Артефакты мигания и их влияние на зависимость амплитуды вызванных потенциалов от интенсивности стимула. — Физиология человека, 1981, т. 7, № 6, с. 1082—1086.
5. Кочубей Б. И., Рутман Э. М. Динамика психофизиологических показателей в связи с отношением к эксперименту и оценкой «неприятности» стимула. — Новые исследования в психологии, 1982, № 2.
6. Латаш Л. П. Гипоталамус, приспособительная активность и электроэнцефалограмма.— М., 1968. —295 с.
7. Небылицын В. Д. Психофизиологические исследования индивидуальных различий.— М., 1976. —336 с.
8. Соколов Е. Н. Нервная модель стимула и ориентировочный рефлекс. — Вопросы психологии, 1960, № 4, с. 61—72.
9. Урбах В. Ю. Биометрические методы. — М., 1964. —515 с.
10. Фонсова Н. А., Аронс Е. К., Васильева В. М. Электроэнцефалографические корреляты точности воспроизведения временного интервала у человека. — Биологические науки, 1981, № 12.
11. Черниговский В. Н., Мусящикова С. С., Синяя М. С., Мокрушин А. А. Привыкание в висцеральных системах. — Л., 1980. — 242 с.
12. Alexander R., Epstein S. Reactivity to heteromodal stimulation as a function of stimulus intensity and inner arousal.— Psychophysiol., 1978, v. 15, p. 387—393.
13. Averill J. R., Rosenn M. Vigilant and nonvigilant strategies and psychophysiological stress reactions during the anticipation of electric shock. —J. Pers. Soc. Psychol., 1972, v. 23, N 1, p. 128—141.
14. Averill J. R. Personal control over aversive stimuli and its relationship to stress. — Psychol. Bull., 1973, v. 80, p. 286—303.
15. Averill J. R. A selective review of cognitive and behavioral factors involved in the regulation of stress. — In: Depue R. A. (ed.) The psychobiology of the depressive disorders. N. Y.: Academic Press, 1979.
16. Birchall P. M. A., Claridge G. S. Augmenting— reducing of the visual evoked potential as a function of changes in skin conductance level. — Psychophysiol., 1979, v. 16, N 3, p. 482—490.
17. Brown W. R. Conceptions of perceptual defence. Cambridge, 1961.
18. Buchsbaum M. Self-regulation of stimulus intensity. — In: Schwartz G., Shapiro D. (eds.) Consciousness and self-regulation N.Y.: Plenum Press, 1976, p. 101—135.
19. Caroll D. Physiological response to relevant and irrelevant stimuli in a simple reaction time situation. — In: Man under stress. Proc. 5-th Ann. Conf. Univ. Adelaide, 1972.L, 1974, p. 21—28.
20. Gale A., Coles M., Kline P., Penfold V. Extraversion — intraversion and the EEG: Basal and response measures during habituation of the orienting reflex. — Brit. J. Phychol., 1971, v. 62, N 4, p. 533—544.
21. Graham K. Habituation and dishabitua-tion of responses innervated by the autonomic nervous system. — In: Peeke H. V., Hertz M. J. (eds.) Habituation, v. 1, 1973, N.Y., p. 1163— 1212.
22. Fahrenberg G. Physiological concepts in personality research. — In: Catell R., Dreger R.
101
(eds.) Handbook of modern personality theory. N.Y., 1978.
23. Harris R. M. Conceptual complexity and prefered coping strategies in anticipation of temporally predictable and unpredictable threat— J. Pers. and Soc. Psychol., 1981, v. 41, N 2, p. 380—390.
24. Israel N. R. Leveling-sharpening and the anticipatory cardiac response. — Psychosom. Med., 1969, v. 31, N 6, p. 499—509.
25. Koriat A., Averill J. R., Malmstrom E. J. Individual differences in habituation: Some methodological and conceptual issues. — J. Res. in Personal., 1973, v. 7, N 1, p. 88—101.
26. Lewis N. A developmental study of learning within the. first three years of life: Response decrement to a redundant signal. — Res. Bull. Princeton, N. Y.: Educational testing servis, 1968.
27. Loveless I. Event-related brain potentials in selective response. — Biol. Psychol., 1977, v. 5, p. 135—149.
28. Maltzman I. The orienting reflex and thinking as determiners of conditioning and generalisation to words. — In: Spence K. (ed.) Essays in neobehaviorism. N.Y., 1971, p. 89— 112.
29. Maltzman I., Raskin D. C. Selective orienting and habituation of the GSR as a consequense of overt and covert activity. — Physiol. Psychol., 1979, v. 7, p. 204—248.
30. Ohman A., Frederikson M., Hugdahl N. Orienting and defensive responding in the electrodermal system: Palmar-dorsal differences and recovery rate during conditioning to potentially phobic stimuli.— Psychophysiol., 1978, v. 15, N 2, p. 93—99.
31. Raskin D. C. Attention and arousal,—In: Prokasy W., Raskin D. C. (eds.) Electro-dermal activity in psychological research. N. Y.: Acad. Press, 1973, p. 125—155.
32. Raskin D. C, Kotses H., Bever J. Autonomic indicators of orienting and defensive responses. — J. Exp. PsychoL, 1969, v. 80, p. 429—433.
33. Routtenberg A. The two-arousal hypothesis: Reticular formation and limbic system. — Psychol. Rev., 1968, v. 75, N 1, p. 51—80.
34. Rust J. Habituation and the orienting response in the auditory cortical evoked potential. — Psychophysiol., 1977, v. 14, N 2, p. 123—126.
35. Siddle D. A. T., Heron P. A. Reliability of electrodermal habituation measures under two conditions of stimulus intensity. — J. Res. Pers., 1976, v. 10, N 2, p. 195—200.
36. Tecce J. J., Savingano-Bowman J., Cole J. O. Drug effects on contingent negative variation and eyeblinks: The distraction-arousal hypothesis. — In: Lipton M. A; (ed.) Psychopharmacology: A generation of progress. N. Y.: Raven Press, 1978.
37. Thompson R. F., Spenser W. A. Habituation: A model phenomenon for the study of neuronal substrate of behaviour. — Psychol. Rev., 1966, v. 73, N 1, p. 16—43.
38. Waid W. M. Perceptual preparedness in man: Brief forewarning reduces electrodermal and psychophysical response to noxious stimulation. — Psychophysiol., 1979, v. 16, N 3, p. 215—221.
39. Weber B. A. Habituation and dishabituation of the averaged auditory evoked response.—J Speech Hear. Res., 1970, v. 13, p. 387— 394.
Поступила в редакцию 12.VIII 1982г.
1 Здесь и далее употребляется термин «привыкание» для обозначения снижения ответов возбуждаемой системы при повторении стимуляции без анализа «истинности» привыкания в том смысле, как она определяется в классической работе Томпсона и Спенсера [37] или в недавно вышедшей книге В.Н. Черниговского с соавт. [11]. Возможно, что «истинное» привыкание составляет лишь часть наблюдавшихся нами снижений реакций при повторении стимула. Для части исследованных реакций привычней был бы термин «угашение» (например, угашение КГР), но поскольку термин «угашение» обычно применялся в отношении полностью прекращавшихся реакций, а в нашем исследовании они чаще лишь уменьшались, а не исчезали, мы решили использовать термин «привыкание» для всех случаев.
2 Здесь и далее приняты сокращения: стр. — строчка, ст. — столбик