105
ПСИХОФИЗИОЛОГИЧЕСКИЕ
КОРРЕЛЯТЫ ВОСПРИЯТИЯ ЗРИТЕЛЬНЫХ СТРУКТУР: БЛИЗНЕЦОВОЕ ИССЛЕДОВАНИЕ
Т. Г. ИВОШИНА
При изучении взаимодействия наследственных и средовых влияний в процессе индивидуального развития человека особое значение приобретает метод регистрации вызванных потенциалов (ВП). Индивидуально-своеобразная и относительно устойчивая форма этих реакций дает основание для постановки вопроса об их генотипической обусловленности. Действительно, в целом ряде исследований, выполненных с помощью метода близнецов, были получены экспериментальные доказательства наследственной обусловленности волновой формы и ряда параметров ВП на элементарный зрительный стимул-вспышку [14], [16], [8]. Восприятие такого сигнала близко к элементарному сенсорному процессу — ощущению ахроматического света. В этой связи закономерно возникает вопрос о том, в какой мере наследственные влияния сказываются на конфигурации ВП, регистрируемых в ответ на предъявление различного рода структурированных стимулов, которые в настоящее время широко интегрируются как электроэнцефалографический коррелят процессов приема и переработки визуальной информации [3]. Исследование ВП такого типа в аспекте генетической психофизиологии может дать некоторые представления о соотношении наследственных и средовых влияний в формировании нейрофизиологических механизмов предметного восприятия.
МЕТОДИКА
В данном сообщении будут изложены материалы, касающиеся только самой общей характеристики ВП — их волновой формы. Исследование было проведено методом близнецов. Внутрипарное сходство волновой формы ВП, полученных при предъявлении стимулов разной формы и содержания, сопоставлялось в группах монозиготных и однополых дизиготных близнецов (МЗ, ДЗ)1 по 20 пар в каждой. Возраст 18—25 лет.
Стимулы: диффузный засвет экрана или вспышка (1)2, симметричная геометрическая фигура (2), комбинация букв ДМО (3), хаотический набор элементов или бесструктурное изображение (4), слово ДОМ (5), изображение дома3 (6), шахматный рисунок с ячейкой 20΄ (7) — предъявлялись бинокулярно сериями по 25—30 в каждой. Время экспозиции и паузы варьировало в пределах 1—2 с и 3—5 с соответственно. Постоянная времени 0,3. Стимулы-изображения, выполненные светлыми штриховыми линиями на черном фоне (угловой размер около 4˚), проецировались на черный матовый экран через диапроектор «Кодак». Угловой размер светового пятна равнялся 5 °. Все стимулы были уравнены по освещенности, которая у экрана составляла 10 лк. Расстояние до экрана 90 см. Испытуемые имели нормальное или скорригированное очками зрение. В инструкции, предъявляемой испытуемым перед началом эксперимента, требовалось: внимательно смотреть на экран, фиксируя взглядом центр изображения, не моргать в момент предъявления стимула. По окончании опыта испытуемым предлагалось ответить на ряд вопросов по поводу изображений. ВП регистрировались монополярно в отведениях:
106
Таблица
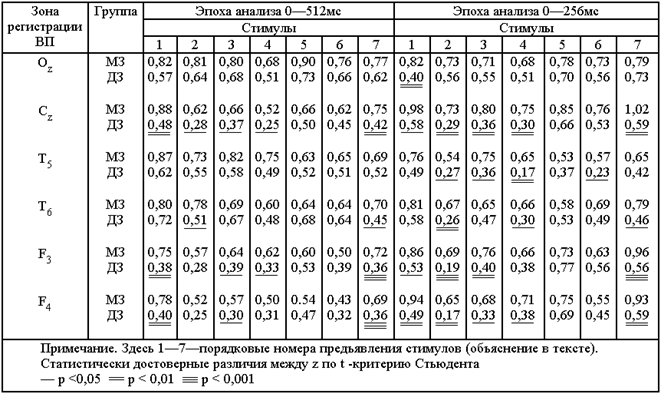
СРЕДНИЕ ЗНАЧЕНИЯ Z, ХАРАКТЕРИЗУЮЩИЕ СТЕПЕНЬ ВНУТРИПАРНОГО СХОДСТВА ВОЛНОВОЙ ФОРМЫ ВП В ГРУППАХ МЗ И ДЗ БЛИЗНЕЦОВ
затылочном (Oz), вертексе (Cz), задне-височных (Т5—Т6) и верхнелобных (F3—F4), в связи с различным эволюционным возрастом зон соответственно обсуждаемой задаче. Референтным служил объединенный ушной электрод. ЭЭГ параллельно регистрировалась на электроэнцефалографе «МЕ-175Е» фирмы «Нихон Кохден» и магнитографе «ТЕАС R-250». Участки ЭЭГ с глазными движениями из обработки исключались. Усреднение ВП производилось на компьютере «АТАС-350» фирмы «Нихон Кохден». Усреднялось по 20 одиночных ответов.
Анализ ВП начинался с преобразования аналоговой формы ВП в цифровую с шагом дискретизации 4 мс. Эпоха анализа составляла 512 мс. Оценка внутрипарного сходства волновой формы ВП производилась в каждой паре МЗ и ДЗ отдельно путем сопоставления соответствующих цифровых рядов с помощью коэффициента линейной корреляции. Сопоставление велось для всего ответа в целом и его раннего фрагмента 0—256 мс. Средние значения коэффициентов корреляции для ВП на каждый из стимулов внутри близнецовых выборок были получены на основе z-преобразования. Поскольку при описании ВП с помощью цифрового ряда и вычислении усредненного коэффициента корреляции точное определени числа степеней свободы затруднительно [15], сравнение групповых данных МЗ и ДЗ близнецов проводилось путем сопоставления соответствующих средних значений z-величин, имеющих нормальное распределение [11]. Достоверность различий определялась с помощью критерия Стьюдента.
РЕЗУЛЬТАТЫ
В табл. представлены средние значения z-величин, характеризующие степень внутрипарного сходства волновой формы ВП в группах МЗ и ДЗ близнецов.
Максимум статистически значимых различий между группами МЗ и ДЗ был получен для ответов в области вертекса; в этом случае волновая форма ВП на все виды стимулов, кроме изображения дома и его вербального обозначения, обнаруживает признаки генотипической обусловленности, причем это справедливо как для всего ответа в целом, так и для его ранней части. Минимум таких различий — в ВП затылочной области; исключение составляет ранний фрагмент ВП на вспышку.
В задневисочной области левого полушария применительно ко всему ответу в целом статистически достоверных различий между МЗ и ДЗ близнецами также не обнаружено. Наряду с этим сопоставление волновой формы ВП в интервале 0—256 мс позволило выявить определенное влияние генотипа в конфигурации ответов на предъявление симметричной геометрической фигуры, бесструктурного изображения, изображения дома, комбинации букв ДМО. В правой задневисочной области волновая форма ВП (и по ответу в целом, и по раннему фрагменту) внутрипарно более сходна у МЗ, чем у ДЗ, в двух случаях: при восприятии симметричной геометрической фигуры и шахматного рисунка. Кроме того, определенное влияние генотипа обнаруживается
107
в рисунке волновой формы раннего фрагмента ВП на бесструктурное изображение.
В верхнелобных отделах обоих полушарий для всего ВП присуще большее сходство МЗ, которое выступило при сравнении ответов на вспышку, комбинацию букв ДМО, шахматный рисунок. При анализе реакций левого полушария к этим стимулам добавляется также бесструктурное изображение. Сопоставление раннего фрагмента ВП в обоих полушариях позволило обнаружить влияние генотипа в рисунке волновой формы ответов на все виды стимулов, кроме изображения дома и его вербального обозначения. Исключение составляет также ранний фрагмент ВП на бесструктурное изображение в левом полушарии.
В целом анализ внутрипарного сходства волновой формы ВП свидетельствует о существовании определенной регионарной специфики проявления генотипических влияний. Конкретно это выражается в относительно высоком уровне наследственной обусловленности волновой формы большинства ответов вертекса и верхнелобных областей обоих полушарий и практически полного отсутствия наследственных влияний в конфигурациях ВП затылочной области.
Отчетливые различия обнаруживаются также в проявлении генотипических влияний в волновой форме ответов на стимулы, различающиеся по форме и содержанию. Относительно высокий уровень наследственных влияний характерен для ответов на элементарный стимул-вспышку и ряд структурированных стимулов, достаточно сложных по форме (симметричной геометрической фигуры, бесструктурного изображения, шахматного рисунка). Эти стимулы, по-видимому, не поддаются однозначной вербализации. В субъективных отчетах испытуемые описывают эти изображения по-разному: «орнамент», «конверт», «скрещивающиеся палки»; «развалившийся домик», «набор черточек», «непонятный рисунок»; «ткань в клеточку», «шахматная доска», «рекламное изображение», «рябь». К этим стимулам примыкает также буквенная композиция ДМО. Практически не сказываются влияния генотипа на волновой форме ВП, зарегистрированных в ответ на предъявление изображения хорошо знакомого объекта и его вербальное обозначение.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Исходной точкой для постановки настоящего исследования явилось представление о возможных различиях в степени генотипической обусловленности разных по эволюционному возрасту и времени дефинитивного оформления в онтогенезе мозговых структур и соответствующих им функций [1].
Иллюстрацией данного положения является еще раз продемонстрированная в настоящем исследовании высокая наследственная обусловленность рисунка волновой формы ВП, регистрируемых в ответ на изменение уровня освещенности. Этот вид перцепции, являясь филогенетически наиболее древним, опосредуется относительно элементарными нейрофизиологическими механизмами [7], [1]. Соответствующие ВП могут быть зарегистрированы даже у недоношенного ребенка, а к концу первого полугодия они приобретают многокомпонентную структуру, достаточно близкую к дефинитивной форме ответа [3].
Более сложную и неоднозначную картину дало исследование роли генотипических влияний в рисунке волновой формы ВП, регистрируемых на стимулы разного вида. Последнее представляется достаточно закономерным, если иметь в виду, что в целом вопрос о путях влияния средовых воздействий на формирование нейрофизиологических механизмов зрительного восприятия очень сложен. Во-первых, противоречивы имеющиеся в настоящее время представления о природе этих механизмов [13], [5], [12]; во-вторых, неясно, какие именно факторы среды оказывают на них наиболее существенное формирующее воздействие. Относительно мало известно также о возрастных изменениях этих механизмов [2], [3]. Существенно, что уже у младенцев могут регистрироваться ВП на самые различные варианты сенсорной стимуляции [21], [25]. Однако эти реакции достаточно далеки от дефинитивной формы ответа и претерпевают весьма существенные изменения в ходе индивидуального развития. При этом, по-видимому, в онтогенезе существует по крайней мере несколько периодов, когда формирующиеся нейрофизиологические механизмы особенно чувствительны к средовым воздействиям [20]. Например, один из первых длится с 3-го до 5-го месяца жизни и связан с формированием механизмов восприятия контура [26]; другой, по-видимому завершающий, приходится на 6—7-й год жизни ребенка и связан с формированием системной организации зрительного восприятия [2]. Приведенные факты достаточно убедительно свидетельствуют о том, что нейрофизиологические механизмы предметного восприятия подвергаются сильным модифицирующим воздействиям. Видимо, поэтому при анализе внутрипарного сходства ВП в группах МЗ и ДЗ, несмотря на более высокий уровень сходства у первых, практически не удалось выявить генотипическую обусловленность рисунка волновой формы большинства ответов, наиболее прямо связанной с процессами зрительного восприятия затылочной зоны. Необходимо, однако, отметить, что волновая форма ВП представляет собой комплексный показатель. Поскольку компонентный состав ВП неоднороден по своему генезу и функциональному значению, можно ожидать, что при дифференцированном анализе параметров отдельных компонентов ВП обнаружится иная картина в проявлении генотипических влияний4. На этом фоне обращает на себя_ внимание тот факт, что в рисунке ВП вертекса и верхнелобных областей обоих полушарий, напротив, генотипические влияния выступают достаточно отчетливо. Последнее, однако, может быть отчасти обусловлено тем, что потенциалы, регистрируемые в этих зонах, достаточно тесно связаны с процессами ориентировочной реакции, общей неспецифической активации, избирательного
108
внимания. Вопрос о том, в какой степени изменения ВП в различных зонах, связанные с разными формами внимания, модально специфичны, остается открытым [28], [23], [18]. Между тем в исследовании [8] было показано, что в ситуации ориентировочной реакции и мобилизации произвольного внимания генотипическая обусловленность рисунка волновой формы ВП на вспышки и в вертексе, и во фронтальной области выступает более отчетливо. Не исключено, что и в настоящем исследовании относительно высокий уровень генетических влияний, зафиксированный для большинства ответов вертекса и верхнелобных областей, фактически связан с актуализацией подкорковых неспецифических активационных влияний. Наряду с этим определенная степень генотипических влияний была выявлена для ответов обеих задневисочных областей. Височные области по некоторым данным имеют прямое отношение к зрительному гнозису, в частности к опознанию зрительных стимулов [10], [17].
Возможно, полученные в работе факты генотипической обусловленности волновой формы некоторых реакций этих областей, в первую очередь ответов на симметричную геометрическую фигуру, бесструктурное изображение, шахматный рисунок, свидетельствуют об определенном влиянии генотипа на их функционирование. Одной из главных операций при восприятии формы объекта является, как известно, выделение контура. Узнавание фигуры происходит на основе объединения информации об отдельных участках ее контура в единый код, который формируется на основе врожденных способностей зрительной системы к восприятию перепадов освещенности и кривизны [6]. Как уже говорилось, критический период, связанный с развитием способности воспринимать контур, длится с 3-го по 5-й месяц жизни [26]. Таким образом, способности такого рода формируются в своей основе достаточно рано. Остается, однако, открытым вопрос о том, каково структурно-функциональное обеспечение этого процесса. Полученные в настоящем исследовании факты свидетельствуют об определенном влиянии генотипа на эти механизмы.
Специальное внимание необходимо уделить результатам внутрипарного сопоставления ВП, зарегистрированных в ответ на вербальный стимул — слово ДОМ и комбинацию букв ДМО. Весьма существенно, что для первого стимула ни в одном из вариантов сопоставления МЗ и ДЗ не было зафиксировано проявлений генотипической обусловленности волновой формы ответа, для второго, напротив, генотипические влияния в рисунке волновой формы ВП были обнаружены в целом ряде случаев. Анализируя этот факт, следует иметь в виду, что при восприятии вербальных стимулов функционирование конкретного сочетания букв в качестве элемента восприятия определяется его произносимостью и положением в слове. При этом испытуемые могут менять размерность элементов в зависимости от целого ряда факторов: тренировки, уровня мотивации, внимания и т. д. [7], [24]. Очевидно, таким образом, что восприятие этих мало различающихся по физическим параметрам стимулов осуществлялось по-разному. По-видимому, различным должно быть и нейрофизиологическое обеспечение этих процессов.
Тэтчер [27], рассматривая проблему восприятия речи, выдвигает представление о двух уровнях нейрофизиологического обеспечения: первичном, сенсорном, отражающем наиболее элементарные параметры речевых стимулов (гласные, согласные звуки, фонемы), и вторичном, отражающем большие и более значимые единицы речи (морфемы, слова, фразы и т. д.). При этом отмечается, что первичный уровень теснее связан с функционированием филогенетически более древних образований, вторичный — с функционированием «специфически человеческих» молодых структур, опосредующих приобретение социального опыта. В онтогенезе латерализация лингвистических функций низшего уровня — фонетического и фонологического — в общих чертах завершается к концу 1-го года жизни. Вопрос о сроках латерализации других лингвистических функций, связанных с синтаксисом, семантикой и включением речевой памяти, остается открытым [22]. В таком контексте представляется закономерным тот факт, что ВП на комбинацию букв ДМО обнаруживают определенную степень генотипической обусловленности, а ВП на слово ДОМ практически не подвержены влияниям генотипа. В том же русле лежат экспериментальные факты [9]: при анализе генотипической обусловленности параметров ЭЭГ был выявлен факт значительно меньшей генотипической детерминированности по сравнению с другими зонами ЭЭГ левой височной зоны. Последнее расценивается как следствие связи этой области с высшими когнитивными процессами, в первую очередь речевыми.
Таким образом, полученные в настоящем исследовании данные о средовой детерминированности конфигурации ВП на хорошо знакомый вербальный стимул представляют собой, видимо, проявление более общей закономерности, в соответствии с которой нейрофизиологические механизмы речевых процессов в своем развитии направляются социальными воздействиями и лишь, возможно, в самых низших своих уровнях сопряжены с актуализацией генетических потенций.
В заключение следует отметить, что в развитии нейрофизиологических механизмов зрительной перцепции достаточно отчетливо прослеживается эволюционно-генетический принцип, в соответствии с которым элементарные перцептивные процессы и опосредующие мозговые механизмы больше зависят от влияния генотипа, нежели относительно молодые длительно формирующиеся в онтогенезе виды перцепции и их структурно-функциональное обеспечение.
ВЫВОДЫ
1. Внутрипарное сопоставление волновой формы зрительных ВП у МЗ и ДЗ близнецов свидетельствует о существовании региональной специфики в проявлении генотипической обусловленности волновой формы ответов на стимулы различного вида, что, по-видимому, связано с различным вкладом этих структур в обеспечение перцептивного процесса.
2. Относительно высокий уровень генотипических влияний характерен для ответов на
109
элементарный стимул — вспышку и ряд структурированных стимулов: симметричную геометрическую фигуру, бесструктурное изображение, шахматный рисунок. Последнее свидетельствует об определенном влиянии генотипа на формирование межиндивидуальной изменчивости нейрофизиологических механизмов этих реакций.
3. Практически не обнаружено влияния генотипа в рисунке волновой формы ВП, регистрируемых в ответ на хорошо знакомый вербальный стимул, что можно расценивать как проявление более общей закономерности, связывающей формирование нейрофизиологических механизмов речевых функций с определяющим воздействием социального опыта.
1. Ата-Мурадова
Ф. А. Развивающийся мозг. Системный анализ.— М., 1980. — 296 с.
2. Бетелева
Т. Г. Формирование системной организации зрительного восприятия в
онтогенезе: Автореф. докт. дис. — М., 1981. — 45 с.
3. Бетелева
Т. Г., Дубровинская Н. В., Фарбер Д. А. Сенсорные механизмы развивающегося
мозга. — М., 1977. — 176 с.
4. Гиндилис
В. М., Феногенова С. А., Животовский Л. А. Некоторые аспекты генетического
анализа полигенных признаков человека на основе семейных корреляций. — В сб.:
Проблемы генетической психофизиологии. М., 1978, с. 196—221.
5. Глезер
В. Д. и др. Зрительное опознание и его нейрофизиологические механизмы. —
Л., 1975. — 269 с.
6. Грановская
Р. М., Березная И. Я., Григорьева А. Н. Восприятие и признаки формы.—М.,
1981. —208 с.
7. Конорски
Ю. Интегративная деятельность мозга. — М., 1970. — 412 с.
8. Марютина
Т. М. О генотипической обусловленности вызванных потенциалов человека. — В
сб.: Проблемы генетической психофизиологии человека. М., 1978, с. 72—93.
9. Мешкова
Т. А. Наследственная обусловленность некоторых параметров
электроэнцефалограммы покоя человека. — В сб.: Проблемы генетической
психофизиологии человека. М., 1978, с. 48—71.
10.
Милнер П. Физиологическая психология. — М., 1973.— 647 с.
11. Фишер
Р. А. Статистические методы для исследователей. — М., 1958. — 262 с.
12. Шевелев
И. А. Нейрофизиологические основы зрительного восприятия форм объектов. — В
сб.: Физиология человека и животных. М., 1976, т. 18.
13. Шехтер
М. С. Психологические проблемы узнавания. — М., 1967. — 220 с.
14. Buchsbaum М. S. Average evoked responses and stimulus intensity in identical and
braternal twins. — Physiol. Psychol., 1974, v. 2, N 3 A.
15. Donchin E. Data analysis techniques in
everage evoked potential reseach. — In: Average evoked potentials: Methods
results and evaluation. Washington, D. C: NASA SP-191, 1969, p. 199—236.
16 Dustman R. E., Schenkenberg Т.,
Lewis E. G. The cerebral evoked potential: Life—span
changes and twin studies. — In: Destmedt J. (ed.) Visual evoked potential in
man: New development.
17. Gross С G., Mishkin M. The neural basis of stimulus equivalence across retinal translation. —
In: Harnad S. et al. (eds.) Lateralization in the nervous system. Academic
Press, 1977, p. 109—122.
18. Harter M. R., Guido W. Attention to pattern
orientation: Negative cortical potentials, reaction time, and the selection
process. — EEG and Clin. Neurophysiol., 1980, v. 49, N 4, p. 461—475.
19. Maurer D., Lewis T. A physiological
explanation of infants early visual development. — Canad. J. Psychol., 1979, v.
33, N 4, p. 232— 252.
20. Mitchell D. S. Effect of early visual
experience on the development of certain perceptual abilities in animals and
man. — In: Walk, Pick H. (eds.) Perception and Experience. Plenum Press, 1978,
p. 37—75.
21. Molfise D. Infant cerebral asymmetry.—
In: Segalowitz S. J., Gruber F. A. (eds.) Language development and neurological
theory. Academic Press, 1977, p. 22—23.
22. Moscovitch M. The development of
lateralization of language function and its relation to cognitive and
linguistic development: A review and some theoretical speculations. — In:
Segalowitz S. J., Gruber F. A. (eds.) Language development and neurological
theory. Academic Press, 1977, p. 194—207.
23. Naatanen R., Michie P. Differents
variants of endogenous negative brain potentials in performance situations: A
reveiw and classification. — In: Lehmann D., Callaway (eds.) Human evoked
potentials applications and problems. N. Y.: Plenum Press, 1979, p, 251—269.
24. Santa J., Santa C, Smith E. Units of
word recognition Evidence for the use multiple units. — Percept, and
Psychophys., 1977, v. 22, N 6, p. 585—591.
25. Schulman-Galambos C., Galambos R.
Cortical responses from adults and infants to complex visual stimuli. EEG and
Clin. Neurophysiol., 1978, v. 45, N 4, p. 425—435.
26. Spekreijse H. Maturation of Contrast EPS
and development of visual <resolution. — Arch. Hal. Biol., 1978, v. 116, p.
358—369.
27. Thatcher R. W. Issues in
neurolingiustics: evoked potential analysis of cognition and language. — In:
Otto D. (ed.) Multidisciplinary perspertives in event-related brain potential
research.
28. Van Voorhis S., Hillyard S. A. Visual
evoked potentials and selective attention to points in space. — Percept, and
Psychophys., 1977, v. 22, N 1, p. 54—62.
Поступила в
редакцию 17. XII. 1982 г.
1 Зиготность близнецов определялась с помощью
анатомо-морфологических, а в ряде случаев дерматоглифических обследований
сотрудниками лаборатории фармакогенетики НИИ по БИХС АН СССР
2 Номера в скобках — порядок предъявления.
3 Изображение дома и
симметричная геометрическая фигура складывались из элементов, составляющих 4-й
стимул.
4 В нашем исследовании
проведен амплитудно-временной анализ отдельных компонентов ВП, но в данной
статье эти результаты не обсуждаются.