107
ИНДИВИДУАЛЬНЫЕ ОСОБЕННОСТИ ПРОСТРАНСТВЕННОЙ СИНХРОНИЗАЦИИ БИОПОТЕНЦИАЛОВ ГОЛОВНОГО МОЗГА В ЗАВИСИМОСТИ ОТ СВОЙСТВА СИЛЫ НЕРВНОЙ СИСТЕМЫ
Е. Р. БЕЛЯЕВА
В исследованиях свойств нервной системы (СНС) широкое распространение получила, электроэнцефалография. Показателями СНС служат амплитудные, частотные и другие характеристики локальной ЭЭГ. До недавнего времени в дифференциальной психофизиологии не использовалась еще одна возможность регистрации ЭЭГ, которая позволяет оценить пространственно-временную, системную организацию мозговых функций. Лишь в последние годы появились публикации о первых работах, включающих такой подход в сферу дифференциальной психофизиологии [1], [2], [3]. Он имеет большое значение для исследования индивидуальных психологических особенностей, так как позволяет сопоставить системно организованную психическую функцию с системными же нейрофизиологическими процессами.
Согласно современным представлениям, именно системное объединение различных отделов мозга обеспечивает его интегративную функцию. Одним из условий такого-объединения является пространственная синхронизация (ПС) биопотенциалов мозга [4]. Установлено, что сходство динамики биоэлектрических процессов является отражением функциональной взаимосвязи пространственно удаленных пунктов коры и что существует зависимость между увеличением общего уровня синхронности в топографически удаленных структурах мозга и повышением их функциональной активности.
Основываясь на концепции М. Н. Ливанова о дистантной синхронизации биопотенциалов пространственно разобщенных пунктов коры как одном из условий их системного взаимодействия, многие исследователи установили связь между особенностями ее организации и спецификой осуществляемой деятельности [3], [5], [6], [7] и др.
Задачей нашей работы явилось исследование индивидуальных особенностей ПС у детей 7—8 лет и решение вопроса о том, отражается ли в них и каким образом свойство силы нервной системы.
МЕТОДИКА
Определение силы нервной системы производилось с помощью двигательной методики В. Д. Небылицына [8], основанной на зависимости времени двигательной реакции От возрастающей интенсивности стимулов. Эта зависимость выражается кривой, крутизна наклона которой является показателем силы — слабости нервной системы по отношению к возбуждению и обозначается коэффициентом b из уравнения регрессии у=а+bх.
Регистрация ЭЭГ осуществлялась на
шестнадцатиканальном чернильнопишущем энцефалографе типа KNG
4756, монополярно в лобных, центральных, теменных, затылочных и височных
областях обоих полушарий, объединенный индифферентный электрод крепился на
мочке уха. Степень ПС оценивалась методом знаковой корреляции, разработанным Т.
П. Хризман [9] и Е. Б. Сологуб [10], который основан на подсчете числа
совпадающих знаков, определяющих однонаправленные изменения попарных ЭЭГ-реализаций
во всех регистрируемых областях. Коэффициенты корреляции (КК) вычислялись по
формуле ![]() . Все значения
КК, находящихся в пределах 0,45—1,0, условно делятся на низкие с величиной
0,45<КК<0,71 и высокие KK ≥
O,71, обозначающие соответственно низкую и высокую
степень ПС биопотенциалов. Поскольку, согласно представлениям ряда авторов [5],
[12], [13], количество именно высоких корреляций отражает степень вовлечения
данного участка-мозга в общую с другими участками деятельность и является
показателем степени активации, мы анализировали только КК ≥ 0,71.
. Все значения
КК, находящихся в пределах 0,45—1,0, условно делятся на низкие с величиной
0,45<КК<0,71 и высокие KK ≥
O,71, обозначающие соответственно низкую и высокую
степень ПС биопотенциалов. Поскольку, согласно представлениям ряда авторов [5],
[12], [13], количество именно высоких корреляций отражает степень вовлечения
данного участка-мозга в общую с другими участками деятельность и является
показателем степени активации, мы анализировали только КК ≥ 0,71.
Показатели ПС изучались в состоянии спокойного бодрствования, при реализации реакции выбора и в ситуации вероятностного прогнозирования. Обе нагрузки реали-зовывались при предъявлении очень близких по структуре стимульных рядов; предполагалось, что это позволит уловить индивидуальные различия, связанные с изменением инструкции.
Для реализации реакции выбора была использована модель случайных событий, состоящая из трех равновероятных сигналов, каждый из которых определял дифференцированный ответ испытуемого: сигнал с Р = 0,7 (светлый квадрат на темном фоне) —
108
двигательную реакцию правой руки, сигнал с Р =,0,2 (шахматный паттерн) — реакцию левой руки, сигнал с Р = 0,1 (контрольная лампочка) — отсутствие двигательной реакции. Сигналы (общее количество 50) предъявлялись тахистоскопически. Каждому сигналу предшествовал предупреждающий стимул. Ответные реакции испытуемого фиксировались по электромиограмме работающих мышц, регистрируемой в полиграфической записи одновременно с ЭЭГ.
Для реализации вероятностного прогнозирования использован ряд предъявлений двух сигналов, следующих друг за другом в случайном порядке с Р = 0,8 и Р = 0,2. Процедура опыта та же, что и в ситуации реакции выбора. Наличие двигательной реакции в промежутке между предупреждающим и основным стимулами свидетельствовало о прогнозе испытуемого. Оценка собственно прогностической деятельности осуществлялась по соотношению частот альтернативных прогнозов как отражению субъективного представления о структуре вероятностной среды.
Анализировались безартефактные отрезки ЭЭГ по 1 с в начале, середине и конце деятельности; для характеристики ПС в целом определялся средний для трех отрезков КК1:
В эксперименте участвовали 30 детей в возрасте 7—8 лет, учащиеся I класса одной из московских школ.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Поскольку наша задача ограничена вопросом
о том, существует ли связь между параметрами силы — слабости и особенностями
ПС, и мы не предполагаем исследовать тип этой связи, анализу будут подвергнуты
только крайние по силе — слабости группы. Они выделялись по критерию ![]() . В одну и другую (т. е. в
группы самых «сильных» и самых «слабых») вошло по 7 человек. Различия между
ними оценивались по непараметрическому критерию серий [14].
. В одну и другую (т. е. в
группы самых «сильных» и самых «слабых») вошло по 7 человек. Различия между
ними оценивались по непараметрическому критерию серий [14].
Сопоставление суперпозиций корреляционных связей между ЭЭГ различных зон выявляет значительные топографические различия ПС у детей с разными значениями коэффициента b (см. рис.).
В состоянии относительного покоя группа «слабых» по сравнению с «сильными» характеризуется значительно большим количеством связей между всеми исследованными областями коры. У «сильных» корреляционные связи концентрируются главным образом в лобно-центральных отделах; у «слабых» к максимуму высоких корреляций в лобно-центральных областях прибавляется еще один фокус их — в нижнетеменных отделах мозга.
Достоверные различия между этими группами обнаруживаются и в процентном отношении реальных КК к максимально возможному их числу: 22% — у «слабых» и 15% —у «сильных» (Р<0,05).
В ситуации реакции выбора у испытуемых двух групп отмечается различный характер изменений в пространственной организации биопотенциалов. Данные суперпозиций свидетельствуют о том, что в группе «сильных» фокусы максимальной активности отмечаются между симметричными точками лобной и центральной областей обоих полушарий, а также в теменно-затылочных областях коры, как внутри-, так и межполушарно.
Для пространственно-временной организации биоэлектрической активности мозга «слабых» характерно наличие корреляционных связей максимально удаленных областей коры, что, согласно данным специальной литературы [4], [15], характеризует высокий уровень неспецифической активации коры головного мозга. Общее количество корреляционных связей в данной группе значительно выше, чем у «сильных». Фокусы максимальной активности отмечаются в височных областях обоих полушарий, в теменно-затылочном и лобно-центральном отделах коры головного мозга.
В ситуации вероятностного прогнозирования существенных различий в паттернах ПС между указанными группами не обнаружено. Суперпозирование высоких корреляционных связей свидетельствует, что в группе «сильных» четкая локализация КК отмечается между симметричными точками лобной и теменной коры двух полушарий, между центрально-теменным и теменно-затылочным отделам» правого полушария, а также в височном отделе левого полушария. Пространственное распределение КК в группе «слабых» в основном повторяет картину дистантной синхронизации биопотенциалов «сильных»; общее количество связей у них также практически одинаково (17 и 18% соответственно, р > 0,05).
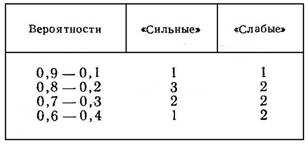
Анализ прогностической деятельности детей с различными показателями силы нервной системы позволил установить следующее распределение стратегий (табл. 1).
109
Все изложенное свидетельствует о том, что в состоянии спокойного бодрствования и в ситуации реакции выбора наблюдаются межгрупповые различия пространственно-временной организации биоэлектрической активности мозга, соотносимые с различиями по силе — слабости нервной системы. Сопоставление картины ПС биопотенциалов у испытуемых двух групп указывает на различную степень генерализованности процесса ПС. Группа «сильных» по сравнению со «слабыми» характеризуется более четкой локальностью и избирательностью формирования фокусов ПС; вместе с тем у «слабых» выраженность процесса общекорковой синхронизации существенно больше.
Таблица 1
КОЛИЧЕСТВО ДЕТЕЙ, ПРОГНОЗИРУЮЩИХ
С РАЗНЫМИ ВЕРОЯТНОСТЯМИ
Согласно современным представлениям, в диффузном повышении синхронизации биопотенциалов отражаются процессы неспецифической активации [15], [16], а в образовании локальных фокусов высокосинхронизированной электрической активности — механизмы избирательного активирования [17]. Исходя из этого, можно полагать, что у обследованных нами детей, различающихся по силе нервной системы, степень участия механизмов локальной и диффузной активации различна. Как известно, лобным областям принадлежит ведущая роль в системе регуляции и контроля мозговой активности ([18] и др.). С этим фактом, по-видимому, связаны полученные нами различия в ПС биопотенциалов лобных областей с остальными корковыми зонами у «сильных». Наше предположение о неодинаковом участии механизмов локальной и диффузной активации у испытуемых с различными типами нервной системы подтверждается
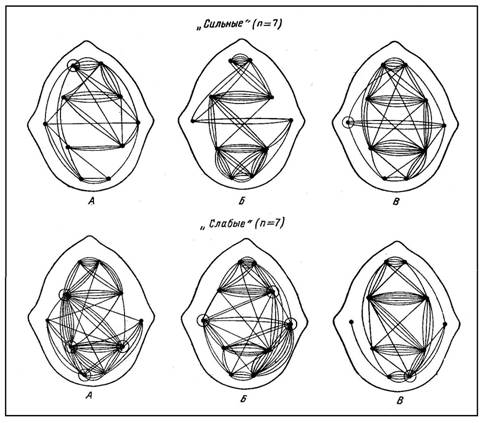
Рис. 1. Суперпозиция высоких межцентральных связей в группах «сильных» и «слабых». А — покой, Б — реакция выбора, В — вероятностный прогноз. Кружками отмечены позиции, в которых количество высоких связей надежно больше, чем в другой группе.
110
данными Д. Грея [19], который отмечает, что параметр силы нервной системы представляет собой показатель уровня активации, т. е. что слабая нервная система — это система более высокого уровня активации, чем сильная.
Таким образом, выраженность генерализованной синхронизации биопотенциалов является основным отличием пространственно-временной организации ЭЭГ у испытуемых с различной силой нервной системы.
В ситуации реакции выбора дети с различными показателями силы нервной системы одинаково справились с заданием. Ни по скорости, ни по количеству ошибок статистически надежных различий между ними не обнаружено. Согласно данным специальной литературы, во всех массовых профессиях, не говоря уже о такой простой деятельности, как двигательная реакция выбора, в равной мере успешные результаты достигаются испытуемыми с различными свойствами нервной системы благодаря индивидуальной стратегии деятельности, компенсирующей особенности нервной системы [20], [21]. Наши данные позволяют предположить, что индивидуальный тип организации активности функционально задействованных корковых областей как раз и лежит в основе индивидуального стиля деятельности, обеспечивающего одинаково успешные результаты работы всех испытуемых.
Основные отличия в локализации фокусов высокосинхронизированной электрической активности заключаются в том, что более высокая связь ЭЭГ височных областей обоих полушарий с ЭЭГ центральных, теменных и затылочных областей отмечается в группе «слабых». Согласно ряду клинических наблюдений [22], [23], [24], височные области имеют отношение к реализации сложных мнестических процессов, процессов опознания, анализа и синтеза предъявляемых сигналов. По-видимому, достижение одинаковых результатов испытуемыми двух групп связано с формированием качественно различных функциональных систем.
В отличие от реакции выбора, в ситуации вероятностного прогнозирования общее: количество связей у испытуемых с разным уровнем силы нервной системы практически одинаково. Объяснением этому феномену может, на наш взгляд, служить эффект обучения: в ряде работ показано нивелирование индивидуальных различий в результате тренировки [20], [25]. Отсутствие таковых сказалось, очевидно, и в паттернах ПС.
Принципиально сходный характер деятельности испытуемых с различной силой нервной системы в условиях вероятностной среды обнаруживается в том, что в группах «сильных» и «слабых» распределение детей, прогнозирующих с разными вероятностями, примерно одинаково.
Таким образом, в паттернах пространственно-временной организации биоэлектрической активности мозга могут отражаться основные свойства нервной системы. В наших экспериментальных условиях особенности ПС, коррелирующие с силой нервной системы, проявились лишь в покое и в реакции выбора. При этом они не коррелируют с успешностью деятельности, что подчеркивает отражение в ПС именно динамической черты поведения — индивидуальной стратегии деятельности.
1. Бодунов М. В. О связи интегративных ЭЭГ параметров с формально-динамическими проявлениями активности человека. — Физиология человека, 1977, №3, с. 394—403.
2. Макаренко С. Л. Пространственно-временная организация электрической активности мозга подростков при сенсорных нагрузках и вероятностном прогнозировании. Автореф. канд. дис. — М., 1979. — 17 с.
3. Русалов В. М. Биологические основы индивидуально-психологических различий. — М., 1979. — 351 с.
4. Ливанов М. Н. Пространственная организация процессов головного мозга. — М., 1972. — 181 с.
5. Хризман Т. П. Развитие функций мозга ребенка; электроэнцефалографические исследования. — Л., 1978.—126 с.
6. Шеповальников А. Н., Цицерошин М., Я., Апанасионок В. С. Формирование биопотенциального поля мозга человека. — Л., 1979. — 162 с.
7. Свидерская Н. Е., Ушакова Т. Н., Шустова Л: А. Электрофизиологическое исследование рече-мыслительного процесса. — Физиология человека, 1980, т. 6, № 3, с. 402—409.
8. Небылицын В. Д. Основные свойства нервной системы человека. — М., 1966. — 383 с.
9. Хризман Т. П. Движения ребенка и электрическая активность мозга. — М., 1973.—187 с.
10. Сологуб Е. Б. Автокорреляционный и кросскорреляционный анализ «меченых» ритмов в. ЭЭГ человека при мышечной деятельности. — Физиологический журнал СССР им. И. М. Сеченова, 1964, т. 50, № 6, с. 681—689.
11. Бернштейн С. Н. Теория вероятности. — 4-е изд., доп. — М.; .Л., 1946.— 556 с.
111
12. Лазарев В. В., Свидерская И. Е., Хамская Е. Д. Дистантная синхронизация биопотенциалов при выполнении произвольных движений. — В сб.: Психологические исследования. М., 1977, с.,71—78.
13. Лазарев В. В., Свидерская Н. Е., Хамская Е. Д. Пространственная синхронизация биопотенциалов при интеллектуальном напряжении в норме и у больных с локальными поражениями мозга. — В кн.: Проблемы нейропсихологии. М., 1977, с. 92—103.
14. Урбах В. Ю. Математическая статистика для биологов и медиков. — М., 1963, с. 311.
15. Хамская. Е. Д. Системные изменения биоэлектрической активности мозга как нейрофизиологическая основа психических процессов. — В кн.: Естественнонаучные основы психологии. М., 1978, с. 234.—253.
16. Хомская Е. Д. Общие и локальные изменения биоэлектрической активности мозга во время психической деятельности. — Физиология человека, 1976, т. 2, № 3, с. 372—386.
17. Фарбер Д. А. Системная организация интегративной деятельности мозга в онтогенезе ребенка. — Физиология человека, 1979, т. 5, № 8, с. 516—527.
18. Лурия А. Р. Основы нейропсихологии. — М., 1973. — 372 с.
19. Грей Д. А. Сила нервной системы, интроверсия — экстраверсия, условные рефлексы и реакция активации. — Вопросы психологии, 1968, № 3, с. 77.
20. Климов Е. А. Индивидуальный стиль деятельности в зависимости от типологических свойств нервной системы. — Казань, 1969. — 278 с.
21. Мерлин В. С. Проблемы интегрального исследования индивидуальности человека. — Психологический журнал, 1980, т. 1, № 1, с. 58—71.
22. Кок Е. П. Зрительные агнозии. — Л., 1967. — 223 с.
23. Прибрам К. Языки мозга: экспериментальные парадоксы и принципы нейрофизиологии. — М., 1975. — 464 с.
24. Брагина Н. Н., Доброхотова Т. А. Функциональные асимметрии человека. — М., 1981. — 287 с.
25. Акимова М. К. Изучение индивидуальных различий по интеллекту. — Вопросы психологии, 1977, № 2, с. 175—185.