74
О ВРЕМЕННОЙ СТРУКТУРЕ СЕНСОМОТОРНЫХ РЕАКЦИЙ ЧЕЛОВЕКА
Т. А. ПАНТЕЛЕЕВА
В психологии одной из наиболее распространенных экспериментальных моделей деятельности человека является сенсомоторное реагирование. При этом в большинстве случаев установление искомых закономерностей осуществляется через анализ изменений времени реакций (BP), отсчитываемого от момента подачи стимула до реализации ответного движения. Вместе с тем известно, что сенсомоторные реакции имеют сложную временную структуру. Целостная реакция предполагает восприятие сенсомоторного стимула, его анализ, принятие решения, моторный ответ и т. д. Это, в свою очередь, заставляет полагать, что выявление динамики только BP в целом, без учета специфических изменений отдельных временных параметров, недостаточно информативно.
В литературе по сенсомоторике человека имеется группа работ, авторы которых выделяют в BP и подвергают отдельному анализу два параметра: премоторный, связанный преимущественно с восприятием и анализом поступающей стимуляции, и моторный, связанный с реализацией движения. Одними экспериментаторами регистрация двигательных реакций осуществляется только по их внешнему проявлению и в «премоторное» время включается период от подачи стимула до начала ответного движения (латентный период — ЛП), в «моторное» — время осуществления самого движения (ВД) [3], [7], [8], [13], [15], [16], [17], [19], [20], [21], [23], [24], [25]. Другие регистрируют двигательные реакции одновременно по их внешнему и ЭМГ-проявлениям. В этих случаях к «премоторному» времени относят период от подачи стимула до появления первых токов действия в мышце, связанной с реализацией движения ЛПЭМГ, а к «моторному» — период от начала ЭМГ до начала регистрируемого движения ЛП—ЛПЭМГ [10], [11], [27], [28] или до момента завершения последнего BP—ЛПЭМГ [12], [22], [26].
Проведенная в указанных работах оценка изменчивости премоторного и моторного параметров показала несхожий характер их изменений в процессе тренировки [3], [8], при смене мотивации [28], сенсорной психологической установки на моторную [13], эмоционального состояния [22], при утомлении [12], [22], [26], при изменении таких характеристик сенсорной стимуляции, как модальность [24] и длительность предпериодов [10], [11], [25], [28], при увеличении трудности сенсомоторного задания [15], [16], [21], [22], [25], при фармакологическом воздействии [7], [8], в онтогенезе [10], [11], [15], [17], [191, [20], [27], [28].
Анализ этих исследований показывает следующее. Чувствительным к изменению длительности предпериодов и модальности стимуляции оказалось только «премоторное» время. Повышение мотивации за счет введения электростимуляции, смена сенсорной установки на моторную,
75
реакция испуга вызвали изменения также в премоторном параметре. Повышение требований к моторному звену (переход по условиям эксперимента от более простых координации к более сложным) сказывалось в увеличении «моторного» времени. В условиях мышечного утомления, по данным одних авторов [22], [26], возрастала величина моторного параметра, других [12] — премоторного. Влияние тренировки оказалось неоднозначным: в одних случаях изменялось только «премоторное» время, в других — только «моторное», в третьих — и то и другое, причем изменения выражались как в увеличении, так и в уменьшении их значений. Правда, были получены факты, говорящие о том, что на динамику этих параметров в ходе тренировки влияет уровень их тренированности и профессиональная специализация испытуемых [8]. Установлено сокращение «премоторного» времени под влиянием брома, сокращение «моторного» и рост «премоторного» под влиянием кофеина [7], [8]. Общая тенденция, выявляющаяся в онтогенетических исследованиях обоих параметров, — уменьшение их величин до определенного возраста с последующим увеличением. Однако период, в который отмечается последовательное уменьшение, длиннее у премоторного (до 19—20 лет) и короче у моторного (до 15 лет). Период стабилизации более длителен у моторного параметра. Возрастное увеличение «премоторного» и «моторного» времени более интенсивно выражено у последнего. Оно начинает проявляться раньше у «премоторного» в группе мужчин (с 19—20 лет) и у «моторного» в группе женщин (с 21 года). При сопоставлении результатов возрастных исследований временных параметров BP, в которых «моторное» время состояло из ВД [15], [19], [20] и из ЛП—ЛПЭМГ [10], [11], [27], [28], обнаружено, что в период от 19 до 86 лет наблюдается рост величины и ВД и ЛП—ЛПЭМГ, однако у первого параметра он выражен в три раза сильнее, чем у второго: величина ВД за указанный период изменяется на 40 %, а величина ЛП—ЛПЭМГ — только на 12 %. Анализ этих параметров в разных половозрастных группах выявил половые различия в величине и возрастной динамике только ВД. Возможно, данные параметры являются независимыми составляющими «моторного» времени.
Надо сказать, что большинство указанных работ выполнено на достаточно репрезентативных выборках. В тех же случаях, когда число испытуемых было не очень большим, исследователи компенсировали этот недостаток большим числом измерений [12], [21], [25].
Приведенные факты свидетельствуют о том, что выделение в BP по меньшей мере двух составляющих — «премоторного» и «моторного» времени — уже дает ясное представление о сложности временной структуры сенсомоторных реакций и функциональных взаимоотношений ее составляющих. Они позволяют заключить, что изучение динамики BP должно проводиться с учетом динамики отдельных временных параметров, входящих в BP, и требуют также соотнесения BP с каждым из параметров. В задачу настоящей работы входил анализ временной структуры сенсомоторных реакций: а) при разных интенсивностях сенсорной стимуляции (при выявлении «закона силы»), б) на разных этапах формирования и переделки автоматизированного навыка реакции выбора, в) при разной сложности сенсорной информации.
Известно, что BP уменьшается при увеличении интенсивности стимуляции в процессе тренировки и возрастает при переходе от простой; реакции к реакции выбора и после переделки (после смены сигнальных значений стимулов). Выявление индивидуальной динамики BP при смене интенсивностей стимулов и при переделке широко используется в дифференциальной психофизиологии, в психологии труда и спорта для диагностики силы нервной системы по ее нижнему порогу — уровню чувствительности и для диагностики подвижности нервных процессов
76
[5], [9]. Однако сопоставление изменений отдельных составляющих BP и соотнесение каждой из них с BP в условиях выявления «закона силы» и переделки не производились. Не проводился такой анализ и в разные периоды сформированности навыка, хотя необходимость его очевидна, если учесть обусловленность изменений премоторного и моторного параметров в процессе тренировки исходным уровнем тренированности последних [8]. В упоминавшихся исследованиях динамики «премоторного» и «моторного» времени при повышении трудности сенсомоторного задания имело место повышение требований только к моторному звену реакции. Поэтому в задачу нашей работы и был включен анализ динамики отдельных временных параметров BP при повышении требований к сенсорному звену.
МЕТОДИКА
Испытуемый находился в кресле полулежа с вытянутой вдоль тела правой рукой, повернутой ладонью вверх. Кончиками пальцев с тыльной стороны кисти руки он нажимал на кнопку, находящуюся на правом колене, а по звуковому сигналу, подававшемуся в наушники, должен был как можно быстрее согнуть правое предплечье в локтевом суставе и нажать ладонью на кнопку, находящуюся возле его плеча. За 2 с до подачи звука предъявлялся предупредительный световой сигнал. Электронные счетчики регистрировали (в мс): 1) время от момента подачи звукового стимула до начала движения — латентный период сенсомоторной реакции ЛП; 2) время движения сгибания предплечья ВД. Параметры ЛП и ВД составляли время реакции от момента подачи раздражителя до конца движения BP. Кроме того, на четырехканальном электроэнцефалографе регистрировалась суммарная ЭМГ двуглавой мышцы, несущей функцию сгибателя. Это позволило выделить: 1) латентный период появления электрической активности в мышце ЛПЭМГ; 2) время опережения ЭМГ-реакцией начала двигательного ответа ВО = ЛП—ЛПЭМГ. Оба показателя — в мс.
Таким образом было выделено пять временных параметров: ЛП, ВД, ЛПЭМГ, ВО, BP, причем ЛП = ЛПЭМГ +ВО, ВР = ЛПЭМГ + ВО + ВД, где ЛПЭМГ — «премоторное» время, ВО и ВД — компоненты «моторного» времени (рис. 1).

Регистрация простой реакции осуществлялась в двух методических вариантах. В первом варианте испытуемым предъявляли звуки частотой 1000 Гц и 4 интенсивностей: 40, 60, 80 и 100 дБ над уровнем 0,0002 бара, поступавшие пачками 13 раз подряд каждый в порядке возрастания интенсивности. Во втором — стимулами являлись два звука частотой 1000 и 500 Гц и громкостью 60 дБ над индивидуальным абсолютным слуховым порогом на тон данной частоты. Каждый из этих звуков предъявлялся 10 раз подряд: сначала — 1000, затем — 500 Гц. Показателями простой сенсомоторной реакции являлись средние величины ЛП, ВД, ЛПЭМГ, ВО, BP на каждый звуковой стимул. Для первого варианта производилась оценка
77
выраженности «закона силы» в каждом из параметров: вычислялись коэффициенты В из уравнения регрессии Y=A + BX, описывающего зависимость величины того или иного параметра от интенсивности стимуляции.
Регистрация реакции выбора. Стимулами являлись звуки, предъявлявшиеся во втором варианте регистрации простой реакции. Сначала у испытуемых вырабатывали навык реакции выбора, когда звук 1000 Гц являлся положительным, а 500 Гц — тормозным. После выработки навыка по предварительной инструкции осуществлялась переделка значений этих стимулов: звук 1000 Гц становился тормозным, 500 Гц — положительным. Испытуемый работал до тех пор, пока у него не сформировывался новый навык реакции выбора. Критерием образования автоматизированного навыка в обоих случаях являлось достижение плато: 20 относительно стабильных по величине ЛП (±20%) реакций на положительный стимул при отсутствии ошибок реакций на тормозной. Ошибками считались возникающие в ответ на тормозной сигнал не только двигательные реакции, но и ЭМГ-реакции двуглавой мышцы, не сопровождающиеся движением. Показателями сенсомоторной реакции выбора являлись средние величины ЛП, ВД, ЛПЭМГ, ВО, BP на этапе врабатывания (характеризуется нестабильной величиной ЛП и наличием обеих групп ошибок) и этапе автоматизированного навыка (или этапе автоматизации) до и после переделки.
Таким образом, в использованном нами методическом варианте биомеханическая структура движения в простой реакции и в реакции выбора оставалась неизменной. Менялась только структура сенсорной стимуляции: интенсивность стимулов и трудность сенсорного задания (при переходе от простой реакции к реакции выбора, в процессе формирования автоматизированного навыка реакции выбора и при смене сигнальных значений стимулов).
Эксперимент с каждым испытуемым проводился в следующем порядке: сначала у него регистрировали простую реакцию на звуки одной частоты и разных интенсивностей, затем — простую реакцию на звуки одной интенсивности и разных частот, после чего переходили к формированию у него навыка реакции выбора с последующей переделкой сформированного навыка. Выполнив одно задание, испытуемый переходил к выполнению следующего.
Данная работа являлась составной частью более общей, посвященной исследованию генетической обусловленности сенсомоторных реакций человека методом близнецов. Поэтому в наших экспериментах участвовали близнецы 18—28 лет, 20 монозиготных (МЗ) и 20 однополых дизиготных (ДЗ) пар.
Для определения статистических связей между выделенными показателями применялось вычисление коэффициентов ранговой корреляции Спирмена. Все испытуемые были разделены на 4 выборки по 20 человек в каждой (по две выборки МЗ и ДЗ) так, чтобы один близнец пары оказался в одной выборке, а другой — в другой. В результате были получены 4 выборки: МЗ1 МЗ2, ДЗ1 ДЗ2. Распределение близнецов происходило случайным образом. Подсчет интеркорреляций производился для каждой из выборок в отдельности. Такая обработка определялась следующими соображениями. Объединение близнецов в одну выборку или две (МЗ и ДЗ) для выявления статистических связей между показателями, на наш взгляд, не вполне корректно. Для определения этих связей, согласно требованиям статистики, статистическая выборка должна включать в себя набор случайных и, главное, не связанных друг с другом элементов. Когда лее мы имеем дело с выборкой, состоящей из пар близнецов, т. е. из людей, имеющих полностью (МЗ) или частично (Д3) одинаковый набор генов, указанное выше условие уже не соблюдается.
78
Выявляемые в такой выборке связи могут быть обусловлены какими-то специфическими особенностями самой выборки. Например, особенностями, детерминирующими ту или иную степень внутрипарного сходства близнецов по сопоставляемым показателям (генетическими, средовыми в широком смысле слова и в более узком: особенностями так называемой близнецовой среды в паре и т. д.). Разделение близнецов на четыре упомянутые выборки более удовлетворяло, таким образом, требованию независимости входящих в эти выборки элементов. Кроме того, такое разделение позволило выделить наиболее надежные коэффициенты корреляций, к которым мы относили статистически значимые коэффициенты, устойчиво повторяющиеся во всех четырех выборках [4]. Значимые коэффициенты считались совпадающими, если они соответствовали уровню р<0,05 и выше и были одного знака (при р<0,05; коэффициент ранговой корреляции для каждой из выборок равен 0,456). Анализ статистических связей между выделенными показателями мы будем проводить именно по этим коэффициентам.
РЕЗУЛЬТАТЫ
Сначала рассмотрим динамику премоторного (ЛПЭМГ) и моторных (ВО, ВД) параметров в применявшихся испытаниях (рис. 2).
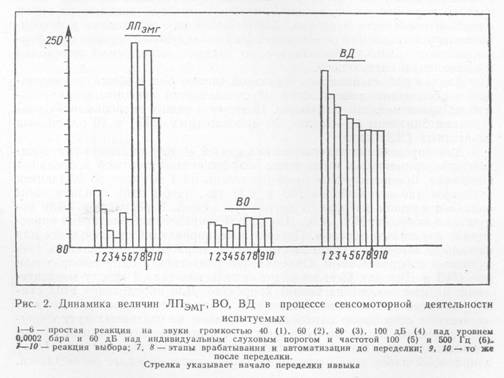
При повышении интенсивности звуков от 40 до 100 дБ над уровнем 0,0002 бара наблюдается уменьшение величин ЛПЭМГ. ВО и ВД. Смена интенсивности стимуляции со 100 дБ над уровнем 0,0002 бара до 60 дБ над индивидуальным уровнем слухового порога испытуемого приводит к увеличению ЛПЭМГ и ВО до значений, близких к тем, которые эти параметры имели в реакции на звук 60 дБ над уровнем 0,0002 бара. ВД при этом продолжает уменьшаться. Переход от простой реакции к реакции выбора вызывает резкое возрастание ЛПЭМГ и небольшое увеличение ВО. ВД становится меньше. В процессе формирования автоматизированного навыка реакции выбора и переделки этого навыка
79
величины ВД и ВО остаются относительно стабильными, изменяется только ЛПЭМГ, уменьшаясь в ходе тренировки и возрастая при смене сигнальных значений стимулов. Выявленные изменения величин ЛПЭМГ, ВО, ВД статистически значимы (оценка значимости производилась по t-критерию Стьюдента).
В целом эти данные свидетельствуют о разной изменчивости «премоторного» времени и компонентов «моторного».
Теперь обратимся к интеркорреляциям выделенных показателей.
Прежде всего отметим, что все статистически значимые коэффициенты корреляций, совпавшие в выборках МЗ1, МЗ2, ДЗ1 ДЗ2, имеют положительный знак.
Тесные устойчивые, воспроизводящиеся во всех четырех выборках связи выявились: 1) в простой реакции — между величинами BP и ВД, BP и ЛП, ЛП и ЛПЭМГ, а также между ВВР и ВВД, ВЛП и ВЛПэмг; 2) в реакции выбора на этапе врабатывания — между величинами ЛП, и ЛПэмг, ЛП и BP, ЛПЭМГ и BP, на этапе автоматизации — между величинами ЛП и ЛПЭМГ, ЛП и BP, ЛПЭМГ и BP, BP и ВД.
Высокие надежные корреляции (0,7—0,9) получены между одноименными параметрами: а) простой реакции на разные интенсивности; б) одноименных этапов формирования навыка. При сопоставлении временных параметров этапов врабатывания и автоматизации устойчивые статистически значимые связи обнаружены между величинами ВД и между величинами ВО.
Не выявлено надежных, повторяющихся во всех четырех выборках корреляций между ЛПЭМГ, ВО и ВД ни по величине, ни по динамике их величин при смене интенсивности стимуляции (по коэффициентам В).
И последнее — несколько слов о процедуре формирования навыка. Напомним, что в наших экспериментах автоматизированный навык считался сформированным при стабилизации величины ЛП, сопровождавшейся отсутствием как ошибочных движений, так и ошибочных ЭМГ-реакций. Следует, однако, отметить, что типичным методическим приемом выработки сенсомоторного навыка обычно является такой: тренировка производится до стабилизации общего времени реакции или латентного периода при отсутствии только ошибочных движений. Недостаточность опоры на эти критерии как единственные для достижения автоматизации следует из полученных нами данных: у 32,5 % испытуемых на уровне плато, соответствующего критериям стабилизации BP и ЛП и отсутствия ошибочных движений, все еще регистрировались ошибочные ЭМГ-реакции. Последние свидетельствуют о том, что в ответ на стимул из центра к эффектору все-таки идет команда отвечать и лишь повторная коррекция и, очевидно, произвольный контроль прерывают начавшуюся реакцию до ее внешнего проявления. Иначе говоря, этот период в процессе формирования навыка еще нельзя считать действительной автоматизацией последнего.
ОБСУЖДЕНИЕ
Первое, что обращает на себя внимание при анализе результатов,— разная динамика ЛПЭМГ, ВО, ВД, составляющих общее время сенсомоторной реакции.
Наиболее чутким индикатором смены структуры сенсорной стимуляции явилось «премоторное» время. В его динамике нашли отражение и изменение интенсивности звуковых сигналов, и изменение трудности сенсорного задания. Последнее имело место в трех вариантах. Первый: при переходе от простой реакции к реакции выбора, означавшем переход от непосредственного восприятия стимуляции к дифференцировке стимулов, имеющих разное сигнальное значение. Второй: в процессе целенаправленного
80
обучения требуемой схеме дифференцирования вплоть до ее автоматизированности и стереотипности (до и в ходе переделки). Третий: при смене сигнальных значений стимулов, потребовавшей перехода к новой, непривычной еще схеме дифференцирования. Динамика ВО оказалась сходной по своей направленности с динамикой «премоторного» времени в условиях смены звуковых интенсивностей и перехода от простой реакции к реакции выбора. Однако изменения ВО в этих условиях имели значительно меньшую выраженность, чем ЛПЭМГ. Смена же сигнальных значений стимулов и формирование автоматизированного навыка реакции выбора до и в ходе переделки не отразились в величине этого моторного параметра. Динамика ВД была обусловлена, по-видимому, процессом целенаправленного обучения требуемому движению. Его величина последовательно уменьшалась до определенного плато: минимального и относительно стабильного значения. Как следует из рис. 2, тренировка ВД завершилась уже к моменту перехода к реакции выбора.
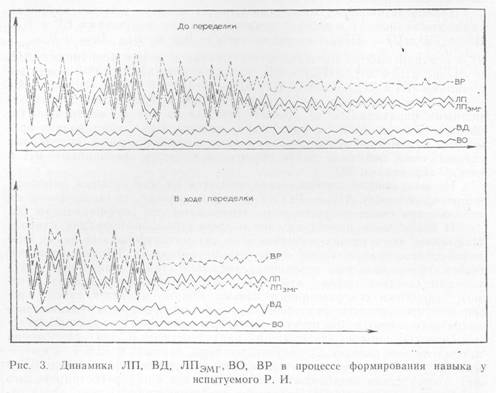
Столь разный характер изменений «премоторного» времени и компонентов «моторного» в целостном сенсомоторном реагировании придает особую значимость сопоставлению этих параметров с общим временем реакции, так как ВР = ЛПЭМГ +ВО + ВД.
Из корреляционного анализа следует, что время простой реакции по величине и по динамике при смене интенсивности стимуляции (по коэффициентам В) обнаружило устойчивую связь только с ВД. В реакции выбора на этапе врабатывания BP показало надежные корреляции с ЛПЭМГ, на этапе автоматизации — с ЛПЭМГ и с ВД.
Такая неоднозначность связи BP с «премоторным» временем и временем движения в простой реакции и реакции выбора на разноименных этапах формирования навыка объясняется, вероятно, тем, что в указанные
81
периоды деятельности испытуемых операциональная роль сенсорного и моторного процессов в целостном сенсомоторном реагировании была разной. Простая реакция отличалась от реакции выбора, во-первых, меньшей информационной сложностью сенсорной стимуляции, во-вторых, непривычностью совершаемых движений. В реакции выбора до и в ходе переделки движения были уже натренированными, но изменялся уровень сложности деятельности по дифференцировке стимулов, имеющих разное сигнальное значение. Этот уровень был максимальным на этапах врабатывания, когда схема дифференцирования была еще непривычной, и минимальным на этапах автоматизации, когда процесс дифференцировки носил автоматизированный и стереотипный характер. Следовательно, чем сложнее и непривычнее характер совершаемых движений и чем проще задача по восприятию и анализу сенсорной стимуляции (что в нашей работе имело место в простой реакции по сравнению с реакцией выбора), тем в большей степени временные характеристики первых будут обусловливать суммарное BP. С другой стороны, чем сложнее деятельность по восприятию сенсорной информации (например, в реакции выбора на обоих выделенных этапах по сравнению с простой реакцией, в реакции выбора в период врабатывания по сравнению с периодом автоматизации), тем в большей степени суммарное BP будет зависеть от «премоторного» времени.
Видимо, при выявлении «закона силы» в простых сенсомоторных реакциях лучше всего оценивать динамику не общего BP (как это делают все исследователи, использующие данный метод для диагностики силы нервной системы), а «премоторного» времени или давать испытуемым возможность освоить требуемые движения. В противном случае изменение BP при изменении интенсивности стимуляции будет обусловлено влиянием не столько смены интенсивностей, сколько особенностями тренировки ВД. Это замечание имеет силу и для методики переделки. В нашей работе стабилизация ВД наступила уже к моменту перехода от простой реакции к реакции выбора. Однако если бы формирование навыка реакции выбора производилось до, а не после регистрации простой реакции, т. е. с самого начала работы испытуемого, то, возможно, BP в период врабатывания до переделки обнаружило бы связь не только с ЛПЭМГ, но и с ВД.
Ни в простой реакции, ни в реакции выбора мы не обнаружили устойчивых связей суммарного BP с ВО. Такой же результат получен и при сопоставлении ВО с ЛП (ЛП = ЛПЭМГ + ВО). Интересно, однако, что, в отличие от BP, латентный период сенсомоторной реакции по своей величине надежно коррелирует с ЛПЭМГ во все периоды деятельности испытуемых: и в простой реакции на все применявшиеся звуковые интенсивности, и в реакции выбора на этапах врабатывания и автоматизации до и после смены сигнальных значений стимулов. ЛП показал устойчивую связь с ЛПЭМГ и по выраженности «закона силы» (коэффициентам В). На рис. 3 представлена типичная динамика всех выделенных временных параметров в процессе формирования навыка реакции выбора до и в ходе переделки. На нем видно, что ЛП и ЛПЭМГ имеют сходную динамику на всем протяжении выработки навыка: и в периоды врабатывания, и в периоды стабилизации.
Другие исследователи сопоставляли ЛПЭМГ, ВО и ЛП только простой реакции при изменении длительности предпериодов [10], [11], [28]. Они установили обусловленность динамики ЛП в этих условиях динамикой ЛПЭМГ. Д. Ботвиник и Л. Томпсон [10], [11] получили высокие коэффициенты корреляций между величинами ЛП и ЛПЭМГ (от 0,87 до 0,96 для n =88), но тесных связей между величинами ЛП и ВО не обнаружили.
82
Результаты нашей работы и указанных выше [10], [11], [28] позволяют заключить, что в условиях изменения структуры сенсорной стимуляции и неизменного характера движений величина и динамика ЛП обусловлены величиной и динамикой преимущественно ЛПЭМГ. По-видимому, использование в данных условиях ЛП в качестве «премоторного» времени, в котором находят отражение прежде всего скоростные характеристики процессов восприятия и анализа поступающей сенсорной стимуляции, возможно. Однако в условиях изменения требований к моторному звену реакции связи ЛП с ЛПЭМГ и с ВО, вероятно, могут оказаться иными. Этот вопрос требует отдельного изучения.
Ряд исследователей при сопоставлении ЛПЭМГ и ВО ([10], [11], [28], [27]), ЛП и ВД ([14], [18] и многие другие), основываясь на фактах разной динамики сравниваемых параметров в онтогенезе и в одних экспериментальных условиях, на отсутствии статистически значимых связей между ними, склоняются к признанию функциональной независимости скоростных характеристик процессов восприятия и моторных процессов в целостном сенсомоторном реагировании.
Этой точке зрения соответствуют результаты и нашей работы: отсутствие хорошо воспроизводимых (повторяющихся в выборках МЗ1, МЗ2, ДЗ1 ДЗ2) статистически значимых связей между ЛПЭМГ, ВО, ВД, между ЛП и ВД, а также разная динамика премоторного и моторных параметров в одних и тех же экспериментальных условиях.
Мы получили также факт функциональной независимости выделенных компонентов «моторного» времени (ВО и ВД). Данные компоненты отражают разные стороны движения: ВО — начало, ВД — длительность. Возможно, в определенных условиях (в том числе и тех, которые имели место в наших экспериментах) временные характеристики процессов, детерминирующих эти стороны моторного акта, оказываются независимыми. По мнению некоторых авторов, период между началом ЭМГ-активности и достижением результата поведенческого акта (у нас: ВО + ВД) связан с функционированием акцептора результатов действия и программы действия [6]. Чувствительность ВО к некоторым характеристикам сенсорной стимуляции, выявленная в нашей работе, возможно, объясняется тем, что функционирование акцептора результатов действия — некоторой афферентной модели, предвосхищающей параметры будущих результатов и определяющей степень совпадения между задуманным и полученным [1], — происходит именно в этот период. Реализация же программы действия осуществляется, вероятно, в основном в течение времени достижения результата.
В заключение коротко остановимся на интеркорреляциях величин одноименных временных параметров реакции выбора. Как уже отмечалось, мы получили надежные и высокие коэффициенты (0,7—0,9) между одноименными параметрами, полученными на одноименных этапах. При сопоставлении разноименных этапов надежные связи выявились только между величинами ВО и между величинами ВД. Эти данные обращают внимание на то, что дифференцировка стимулов, имеющих разное сигнальное значение, в периоды врабатывания и автоматизации обеспечивается, по-видимому, разными механизмами. Основное отличие последних друг от друга — в уровне произвольности процессов саморегуляции, детерминирующих дифференцировку. Этот уровень максимален на этапах врабатывания и минимален — на этапах автоматизации [2].
ЗАКЛЮЧЕНИЕ
Обобщая изложенный в настоящей статье материал, можно сказать следующее.
Регистрация сенсомоторных реакций по их внешнему и ЭМГ-проявлениям
83
позволила выделить в общем времени реакции три временных параметра: 1) латентный период появления ЭМГ-активности в мышце, связанной с реализацией движения ЛПЭМГ; 2) время опережения ЭМГ-реакцией начала движения ВО; 3) время движения ВД. Из них: ЛПЭМГ — «премоторное» время, ВО и ВД — компоненты «моторного» времени. Сопоставление этих параметров друг с другом и с BP при изменении таких характеристик сенсорной стимуляции, как интенсивность и трудность сенсорного задания (при переходе от простой реакции к реакции выбора; в процессе формирования автоматизированного навыка реакции выбора; при переделке сигнальных значений стимулов), и неизменном характере движений (сгибания предплечья) показало следующее.
ЛПЭМГ, ВО и ВД целостной сенсомоторной реакции обнаружили отсутствие устойчивых статистически значимых связей между собой и разную динамику в применявшихся экспериментальных условиях. Наиболее чутким индикатором смены структуры сенсорной стимуляции явилось «премоторное» время. Динамика ВД была обусловлена тренировкой движений. ВО показало небольшую чувствительность к изменению интенсивности стимулов и переходу от простой реакции к реакции выбора. Мы предположили, что, возможно, ВО связано с функционированием акцептора результатов действия, а ВД — с реализацией программы действия.
Суммарное BP обнаружило разную степень связи с каждым из параметров. С ВО оно не показало устойчивых корреляций ни в одном из периодов деятельности испытуемых. Обусловленность BP «премоторным» временем и временем движения зависела от операциональной роли сенсорных и моторных процессов в целостном сенсомоторном реагировании. В простой реакции, когда деятельность по восприятию сенсорной стимуляции характеризовалась минимальным уровнем сложности, но движения были еще непривычными, BP обнаружило зависимость от ВД. В реакции выбора на этапах врабатывания, когда процесс дифференцировки стимулов имел наибольшую сложность и непривычность, а движения были уже натренированными, BP показало устойчивую связь с ЛПЭМГ. В периоды автоматизации реакции выбора, когда и движения и процесс дифференцировки стимулов характеризовались высоким уровнем тренированности, выступила связь BP и с ЛПЭМГ, и с ВД.
Полученные факты, в совокупности с литературными, свидетельствуют о необходимости учета особенностей временной структуры сенсомоторных реакций человека во всех исследованиях, предметом которых являются скоростные характеристики этих реакций.
1. Анохин П. К. Биология и нейрофизиология условного рефлекса. — М., 1968.
2. Бернштейн Н. А. Очерки по физиологии движений и физиологии активности. — М., 1966.
3. Лисенкова В. П. Зависимость времени простой сенсомоторной и дизъюнктивной реакции от фактора тренировки. — В кн.: Экспериментальная и прикладная психология. — Ученые записки Ленинградского университета, 1973, № 368, серия психологических наук, вып. 5, с. 60—67.
4. Мешкова Т. А. Исследование генетической детерминированности различных параметров электроэнцефалографии покоя человека близнецовым методом: Автореф. канд. дис. — М., 1976.
5. Небылицын В. Д. Основные свойства нервной системы человека. — М., 1966.
6. Пашина А. X., Швырков В. Б. О сокращении времени реакции при обучении. — В кн.: Теория функциональных систем в физиологии и психологии. М., 1978, с. 347—357.
7. Пономарев М. Ф. О влиянии кофеина и брома на время латентного и моторного компонентов двигательной реакции человека. — Физиологический журнал СССР, 1958, № 2, с. 97—104.
8. Пономарев М. Ф. О двигательных реакциях в связи с восприятием времени. — Вопросы психологии, 1960, № 3, с. 79—88.
9. Шадрин В. М. Психологические основы формирования двигательного навыка. — Казань, 1978.
84
10. Botwinick J., Thompson L. Premotor and motor components of reaction time. — J. Exp. Psychol., 1966, v. 71, N 1, p. 9—15.
11. Botwinick J., Thompson L. Components of reaction time in relation to age and sex. — J. Genet. Psychol., 1966, v. 108, N 2, p. 175—183.
12. Hanson C., Klimovitch G. Effects of fatigue and laterality on fractionated reaction time. — J. Mot. Behav., 1978, v. 10, N 3, p. 177—184.
13. Christina R. Influence of enforced motor and sensory sets on reaction latency and movement speed. — Res. Quart. Amer. Assoc. Health., 1973, v. 44. N 4, p. 483— 487.
14. Clarke D., Glines D. Relationships of reaction, movement, and completion times to motor, strength, anthropometric and maturity measures of 13-year-old-boys. — Res. Quart. Amer. Assoc. Health., 1962, v. 33, N 2, p. 194—201.
15. Deupree R. H., Simon J. R. Reaction time and movement time as a function of age, stimulus duration, and task difficulty. — Ergonomics, 1963, v. 6, N 4, p. 403—411.
16. Fitts P. M., Peterson J. R. Information capacity of discrete motor responces. — J. Exp. Psychol., 1964, v. 67, N 2, p. 103—112.
17. Fulton C. D., Hubbard A. W. Effect of puberty on reaction and movement times. — Res. Quart. Amer. Assoc. Health., 1975, v. 46, N 3, p. 335—344. 1
18. Нету F. M., Whitley J. D. Relationships between individual differences in strength, speed and mass in an arm movement. — Res. Quart. Amer. Assoc. Health., 1960, v. 31, N 1, p. 24—33.
19. Hodgkins J. Influence of age on the speed of reaction and movement in females. — J. Gerontol., 1962, v. 17, N 4, p. 385—389.
20. Hodgkins J. Reaction time and speed of movement in males and females of various ages. — Res. Quart. Amer. Assoc. Health., 1963, v. 34, N 3, p. 335—343.
21. Jagacinski R. J., Hartzell E. J., Ward S., Bishop K. Fitts'law as a function of system dynamics and target uncertainty. — J. Mot. Behav., 1978, v. 10, N 2, p. 123— 131.
22. Klimovitch G. Startle response and muscular fatigue effects upon fractionated hand grip reaction time. — J. Mot. Behav., 1977, v. 9, N 4, p. 285—292.
23. Laszlo J. I., Livesey J. P. Task complexity, accuracy, and reaction time. — J. Mot. Behav., 1977, v. 9, N 2, p. 171—177.
24. Saari M. J., Pappas B. A. Cardiac cycle phace and movement and reaction times. — Percept, and Mot. Skills, 1976, v. 42, N 3, P 1, p. 767—770.
25. Semjen A., Requin J., Fiori N. The interactive effect of foreperiod duration and responce — movement characteristics upon choice — reaction time in a pointing task. — J. Hum. Mov. Stud., 1978, v. 4, N 2, p. 108—1181.
26. Stull A. G., Kearney J. T. Effects of variable fatigue levels on reaction-time components. — J. Mot. Behav., 1978, v. 10, N 3, p. 223—231.
27. Weiss A. D. The motor components of auditory reaction time as related to age. — American Psychologist, 1956, v. 11, p. 374—383.
28. Weiss A. D. The locus of reaction time change with set., motivation and age. — J. Gerontol., 1965, v. 20, N 1. p. 60—64.