112
ИЗМЕНЕНИЕ ЭЛЕКТРОЭНЦЕФАЛОГРАФИЧЕСКИХ ПРОЯВЛЕНИЙ СВОЙСТВ НЕРВНОЙ СИСТЕМЫ В ЗАВИСИМОСТИ ОТ ПОДКРЕПЛЕНИЯ
Б. Б. ПЫСИН
В учении И.П. Павлова основными показателями свойств общего типа нервной системы были индивидуальные динамические особенности условнорефлекторной деятельности [21]. Когда в лаборатории А. Г. Иванова-Смоленского при изучении детей стали пользоваться различными подкреплениями, то обнаружилось, что у одного и того же ребенка при разных подкреплениях выявляются разные индивидуально-динамические особенности условнорефлекторной деятельности [11], [12], [25], [28]. Поэтому А.Г. Иванов-Смоленский предположил, что наряду с общим типом нервной системы существует парциальный тип, характеризующий индивидуальные свойства той нервной структуры, которой адресовано подкрепление [7].
Различие индивидуально-динамических особенностей условнорефлекторной деятельности в зависимости от подкрепления наблюдалось впоследствии в лаборатории П.С. Купалова [13] и Я. Стреляу [31].
В лаборатории Б.М. Теплова и В.Д. Небылицына установлено, что одни и те же испытуемые обнаруживают различные индивидуально-динамические особенности нервной деятельности в зависимости от модальности условного раздражителя [6], [9], [10], [16], [23]. Поэтому они предположили, что существуют такие парциальные свойства нервной системы, характеризующие нервные структуры, к которым адресован условный раздражитель [18], [19], [26]. Понятие парциальных свойств было применено также к индивидуально-динамическим различиям электроэнцефалографических реакций в зависимости от условного раздражителя [20].
Таким образом, оно приобрело общий смысл и стало широко применяться.
С другой стороны, в свете учения о функциональных системах и соответствующих экспериментальных фактов требует пересмотра истолкование самого феномена парциальности. Согласно П.К. Анохину, тончайшие процессы нервной системы приобретают
113
различный функциональный смысл и значение в зависимости от целостного поведенческого акта, который они обеспечивают [1]. Интеграция отдельных реакций и процессов в целостный приспособительный акт не осуществляется в какой-то специализированной нервной структуре, где происходит «замыкание» нервной связи. Интеграция самых различных приспособительных актов осуществляется в одних и тех же структурах (акцепторе действия). Отсюда вытекает, что индивидуально-динамические различия условнорефлекторной деятельности в зависимости от подкрепления нельзя связывать только со специфическими свойствами подлежащей нервной структуры. При этом, как известно из исследований школы П. К. Анохина, первичная интеграция целостного приспособительного акта осуществляется уже на нейронном уровне. В одних и тех же нейронах сенсорно-соматической коры можно получить условные вызванные потенциалы, различающиеся по динамике, в зависимости от подкрепления [2], [29], [30]. Эти динамические различия точно так же нельзя объяснить парциальными свойствами нервных субстратов, поскольку они проявляются в одном и том же нейроне.
Все это привело к пересмотру первоначального понимания феномена парциальности. Одну из попыток в этом направлении сделал В.М. Русалов [24]. Однако и у него различие в динамике электроэнцефалографических проявлений, в зависимости от модальности условных раздражителей истолковывается как показатель парциального свойства нервной системы.
В.С. Мерлин предложил другую гипотезу для истолкования феномена парциальности [14], [15], основанную на системном подходе. Поскольку свойства нервной системы определяют общее функциональное состояние, то, следовательно, одни и те же индивидуально-динамические особенности определенной реакции могут быть обусловлены разными свойствами нервной системы в зависимости от целостного приспособительного акта, в который они включены. Поэтому феномен парциальности можно объяснить не только тем, что в зависимости от подкрепления изменяются свойства нервной системы, но и тем, что индивидуально-динамические особенности той же реакции становятся выражением другого свойства нервной системы.
Отсюда и возникает основная проблема настоящего исследования. По какому критерию мы в состоянии объективно различать проявление общего и парциального свойства при различных подкреплениях? Если один и тот же показатель при разных подкреплениях коррелирует с показателями одного и того же свойства, но при этом имеет значительно различающуюся величину, эти изменения величины надо истолковывать как проявление парциального свойства нервной системы, связанного с морфологической структурой подкрепляемого очага. Такие данные и были получены в лаборатории А.Г. Иванова-Смоленского.
Если же один и тот же показатель при разных подкреплениях не только изменяется по величине, но и коррелирует с показателями разных свойств нервной системы, эти различия надо истолковывать как проявление разных общих свойств нервной системы. При этом такое сопоставление возможно только в том случае, когда проявления, с которыми мы сопоставляем изучаемый показатель, не связаны с подкреплением, т. е. являются фоновыми.
При таком подходе к проблеме референтным для данного свойства является не какое-либо подкрепление, а показатель, который входит в устойчивый синдром показателей для данного свойства нервной системы.
Излагаемое исследование посвящено проверке изложенной выше гипотезы В. С. Мерлина. С этой целью изучаются индивидуально-динамические особенности одной и той же условной электроэнцефалографической реакции угашения с подкреплением условной депрессии альфа-ритма при двух различных подкреплениях — ориентировочном и оборонительном. Эти особенности сопоставляются с показателями трех свойств нервной системы — силы возбудительного процесса, активированности и лабильности. Если при таком сопоставлении окажется, что динамика угашения условной депрессии альфа-ритма при различном подкреплении коррелирует только с показателями одного и того же свойства нервной системы, то подтвердится истолкование феномена парциальности, предложенное впервые А.Г. Ивановым-Смоленским. Если же при различных подкреплениях будет установлена корреляция с разными свойствами нервной системы, то это соответствует истолкованию феномена парциальности, предложенного В.С. Мерлиным, так как в данном случае при различных подкреплениях динамика угашения условной депрессии альфа-ритма являлась бы показателем не парциальных, а различных общих свойств нервной системы.
МЕТОДИКА
Испытуемыми были 33 студентки в возрасте 18—22 лет. Регистрация биотоков производилась в светоизолированной экранированной камере при помощи 8-канального электроэнцефалографа (тип 4751—2), снабженного анализатором, позволившим выделить из суммарной ЭЭГ полосы частот 1—4, 4—8, 8—13, 13—20, 20—30 кол/с, и интегратором, дававшим индексы электрической энергии по указанным полосам частот. Использовалось биполярное височно-затылочное отведение с правого полушария. После угашения ориентировочной реакции на звук частоты 500 Гц, громкости
114
около 70 дб и длительности 4 с к нему присоединялось подкрепление. В первой серии опытов применялось ориентировочное активирующее подкрепление. На экран, расположенный на расстоянии полутора метров от глаз испытуемого, проецировались сюжетные картинки из диафильмов при помощи фильмоскопа. Подкрепление подавалось автоматически с отставанием от начала звука на 2 с, и в течение 2 с длилось совместно со звуком. Испытуемый получал инструкцию как можно точнее запомнить детали изображения. После выработки условного рефлекса следовала процедура угашения с подкреплением. Предъявлялось 50 сочетаний звука с подкреплением с интервалом 5—10 с между сочетаниями. Для выявления величины условной реакции подкрепление опускалось после 3, 7, 10, 15, 20, 24, 30, 34, 40,- 44, 50 сочетаний. Картинка менялась 2 раза после 10 и 30 сочетаний. За величину условной реакции принималось отношение энергии альфа-ритма во время действия изолированного звука с вычетом времени латентного периода реакции к энергии альфа-ритма за такой же промежуток времени, непосредственно предшествующий подаче звука.
Во второй серии процедура была аналогичной, но подкрепление было другим. Электрокожный раздражитель в виде прямоугольных импульсов частотой 6 имп/с и длительностью 16 мс подавался на указательный палец левой руки. Величина напряжения подбиралась индивидуально — в 2 раза больше порога чувствительности. Испытуемый мог частично избежать тока путем переключения тумблера. Это подкрепление можно квалифицировать как оборонительное.
В двигательной методике измерялось латентное время реакции на звуки разной громкости 45 и 90 дб (модифицированная методика В.Д. Небылицына «закон силы»). Сенсорная методика заключалась в измерении критических частот слияния звуковых щелчков и световых мельканий. Показатели измерялись как среднее из 10 измерений. В результате исследования получены 17 показателей:
1—5. Соответственно энергии ритмов дельта, тета, альфа, бета-1 и бета-2, полученные из 3 м записи фоновой ЭЭГ перед началом опыта при скорости движения ленты 3 см/с.
6—10. Соответственно частоты тех же ритмов. Вычислялись как среднее из 10 измерений в отрезках записи по 1 с из фонового отрезка ЭЭГ.
11. Средний уровень асимметрии единичного колебания ЭЭГ. Был
вычислен как разность восходящих и нисходящих фаз из 30-секупдного отрезка
записи фоновой ЭЭГ при скорости ленты 12 см/с и квантовом шаге 16,7 мс (![]() ).
).
12. Коэффициент асимметрии. Вычислен как отношение восходящей и
нисходящей фазы колебаний (![]() ).
).
13. Критическая частота мельканий (КЧМ).
14. Критическая частота звуковых щелчков (КЧЗ).
15. Отношение среднего времени реакции на сильный звук к среднему времени реакции на слабый звук в двигательной методике.
16. Коэффициент угашения с подкреплением в условно-ориентировочном рефлексе (Куг. ОР). Он вычислялся как отношение величины условной реакции депрессии альфа-ритма в конце опыта к величине ее в начале опыта во время изолированного условного звука. Для каждой картинки он вычислялся отдельно, а затем вычислялось среднее из 3 полученных значений.
17. Коэффициент угашения с подкреплением в инструментально-оборонительном условном рефлексе (Куг. ОБ). Вычислялся аналогичным образом.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
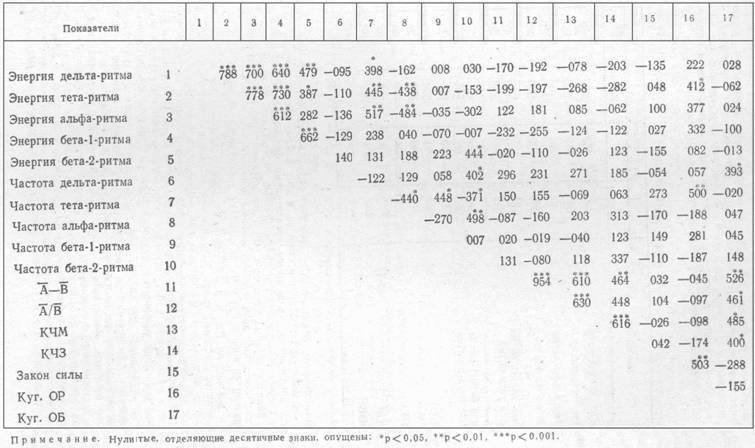
Из корреляционной матрицы (табл. 1) видно, что
коэффициенты угашения с различными подкреплениями не коррелируют друг с другом.
Имеем ли мы здесь то, что обычно называют парциальностью по подкреплению или
коэффициенты являются показателями различных свойств нервной системы? Для
решения этого вопроса обратимся к разбору корреляционных матриц и результатов
факторного анализа. Куг. ОР коррелирует с показателем 15 (закон силы) и с
энергиями тета- и альфа-ритмов, также связываемых с силой нервной системы [4],
[5]. Имеется также значимый коэффициент корреляции с частотой тета-ритма. Куг.
ОБ коррелирует с частотой дельта-ритме, с показателями лабильности КЧМ и КЧЗ и
с показателями ассимметрии ![]() и
и ![]() .
.
Из корреляционной матрицы были извлечены центроидные факторы, и два значимых фактора, выделенных по критерию Фрахтера, подвергнуты вращению. Вращение производилось против часовой стрелки на угол 36°50', и веса вычислены аналитическим способом. Результаты факторного анализа, приведенные в табл. 2, в основном подтверждают корреляционные отношения. После вращения в фактор А с высокими факторными весами вошли энергетические индексы почти всех частотных полос ЭЭГ покоя, за исключением суммарной энергии бета-2-ритма, частота тета-ритма и Куг. ОР. Показатель 15 (закон силы) не набрал значимого веса ни по одному фактору. Это соответствует матрице корреляции: показатель 15 значимо не коррелирует ни с одним показателем фоновой ЭЭГ. Сходный результат был получен В. М. Русаловым. Он также не обнаружил значимой корреляции этого показателя ни с одним
115
Таблица 1
МАТРИЦА РАНГОВЫХ КОРРЕЛЯЦИЙ
116
из интегральных факторов ЭЭГ [24]. Возможно, связь проявления «закона силы» с параметрами фоновой ЭЭГ является более сложной, чем линейная. Возможно, здесь проявляется различие двух функций свойства силы нервной системы — способности клеток выдерживать длительно действующий раздражитель и способности противостоять кратковременному сверхсильному раздражителю.
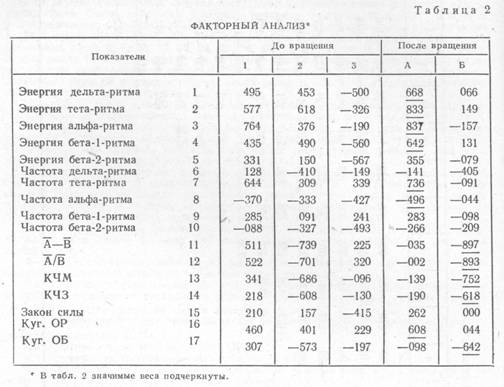
Таким образом, в фактор А, с одной стороны, вошел Куг. ОР, являющийся референтным показателем силы и энергии ритмов дельта и тета, связываемым с силой нервной системы [4], [5]. С другой стороны, в этот фактор вошла энергия альфа-ритма и частота альфа-ритма, являющиеся показателями баланса возбуждения и торможения или уровня реакции активации [4], [17]. Частота тета-ритма также связывается с динамичностью возбуждения и динамичностью торможения, т. е. может быть показателем баланса нервных процессов [18]. Следовательно, фактор А можно интерпретировать как сочетание двух свойств нервной системы — силы возбуждения и уровня активации.
В фактор Б вошел Куг. ОБ, показатели
лабильности нервной системы КЧМ и КЧЗ и показатели ![]() и
и![]() , которые, будучи истолкованы в терминах теории
свойств нервной системы, могут быть использованы как показатели лабильности
[8]. Этот фактор можно интерпретировать как лабильность нервной системы.
, которые, будучи истолкованы в терминах теории
свойств нервной системы, могут быть использованы как показатели лабильности
[8]. Этот фактор можно интерпретировать как лабильность нервной системы.
Таким образом, оказывается, что коэффициенты угашения с различными подкреплениями являются показателями различных свойств нервной системы. ЭЭГ — вариант угашения с подкреплением, как было показано В.Д. Небылицыным [18] и подтверждено в ряде других работ, измеряет силу нервной системы путем многократного проявления сочетаний условного и подкрепляющего раздражителей, которое оказывает истощающее влияние на нервные элементы. Это подтвердилось и в настоящей работе. Куг. ОР является показателем слабости возбудительного процесса. Связь же Куг. ОР с уравновешенностью, возможно, указывает на влияние другого фактора — гипнотического. В.К. Федоров [27] показал, что одной из возможных причин падения величины рефлексов при повторении является гипнотизирующее влияние однообразных раздражителей. Легкое же наступление гипнотического состояния говорит о слабости тормозного процесса. Возможность применения угашения с подкреплением как показателя уравновешенности освещена также в работе Е.С. Роговенко и Е.В. Соколовой [22], проведенной на собаках, где в качестве условного раздражителя использовался звук метронома, а безусловного — пищевое подкрепление. У уравновешенных скорость угашения была быстрее. Что касается связи Куг. ОБ с лабильностью, то можно предположить, что в процессе повторения сочетаний, во мере адаптации
117
к болевому раздражителю, ослабляется доминанта оборонительного очага возбуждения. По Ухтомскому, стойкость доминанты связана с лабильностью — повышенная лабильность ведет к меньшей стойкости. Следовательно, и условный рефлекс, возникший на почве этой доминанты, оказывается менее стойким. Кроме того, на динамику угашения с данным видом подкрепления в процессе длительной однообразной деятельности, возможно, действует фактор утомления. В работе Э.А. Голубевой и Т.К. Василенко [3] показано, что при выполнении определенной деятельности более лабильные испытуемые подвержены более быстрому изменению функционального состояния, в частности нарастанию тормозного процесса. Для разрешения вопроса о том, какой из факторов имеет место или преобладает в данном - случае, требуются дополнительные данные.
Мерилом физиологической силы подкрепления может служить интенсивность реакции, т. е. величина депрессии альфа-ритма. Была подсчитана средняя величина депрессии альфа-ритма для условно-ориентировочного и оборонительного рефлексов из первых трех проб изолированного условного раздражителя. Они соответственно оказались равными 2,039 и 2,415 (меньшее число соответствует большей величине депрессии). Различие средних сказалось значимым на высоком уровне (t= 13,82), р<0,001, т. е. условно-ориентировочная реакция оказалась в среднем более интенсивной, чем условно-оборонительная. Затем испытуемые были разделены на крайние группы с большой и малой величиной депрессии по критерию Х±1/2 σ. Для выделенных групп были сопоставлены средние величины коэффициентов угашения. Оказалось, что для группы с малой величиной депрессии средний Куг. ОБ = 3,157, а для группы с большой величиной депрессии средний Куг. ОБ=3,386. Различие незначимо (t=0,847). Для группы с малой величиной депрессии средний Куг. ОР = 3,266, а для группы с большой величиной депрессии средний Куг. ОР=3,247. Различие также незначимо. Следовательно, коэффициенты угашения не зависят от физиологической силы раздражителя.
ЗАКЛЮЧЕНИЕ
Оказалось, что показатели скорости угашения с различными подкреплениями имеют различный смысл. Если скорость угашения с условно-ориентировочным подкреплением является показателем силы возбудительного процесса и, возможно, уравновешенности, то скорость угашения с оборонительным подкреплением является показателем лабильности. Следовательно, подтверждается гипотеза В.С. Мерлина о том, что в зависимости от различий целостных биологических приспособительных актов одни и те же показатели могут свидетельствовать о различных свойствах типа нервной системы. Этим отнюдь не предрешается вопрос о существовании парциальных свойств нервной системы. Полученные факты говорят лишь о том, что для диагноза парциальности нужны другие критерии, чем применяющиеся сейчас. Что же касается сущности парциальных свойств, функций, которые они выполняют, условий, от которых они зависят, то эти вопросы выходят за рамки настоящего исследования.
1. Анохин П. К. Биология и нейрофизиология условного рефлекса.— М., 1968.— 547 с.
2. Арутюнян С. А. Динамика вызванных потенциалов в процессе выработки пищевого двигательного условного рефлекса. — Журн. высш. нерв, деят., 1970, т. 20, вып. 6, с. 1175—1181.
3. Голубева З. А., Василенко Т. К. Действие кофеина на биоэлектрические показатели при ранних стадиях психического утомления. — В сб.: Типологические особенности высшей нервной деятельности человека. — М., 1965, т. 4, с 203—211.
4. Голубева Э. А., Изюмова С. А., Трубникова Р. С., Печенков В. В. Связь ритмов электроэнцефалограммы с основными свойствами нервной системы. — В сб.: Проблемы дифференциальной психофизиологии. — М., 1974, т. 8, с. 160—174.
5. Гусева Е. П., Шляхта Н. Ф. Некоторые особенности показателей биоэлектрической активности у подростков. — В сб.: Проблемы дифференциальной психофизиологии. — М., 1974, т. 8, с. 199—214.
6. Ермолаева-Томина Л. Б. Концентрированность внимания и сила нервной системы. — В сб.: Типологические особенности высшей нервной деятельности человека. — М., 1959, т. 2, с. 92—106.
7. Иванов-Смоленский А. Г. Экспериментальное исследование высшей нервной деятельности ребенка. — Физиологический журнал СССР, 1935, т. 19, № 1, с. 133—140.
8. Изюмова С. А. Устойчивые характеристики асимметрии как индикаторы свойства лабильности и их связь.с процессами переработки и сохранения информации. — В сб.: Проблемы дифференциальной психофизиологии. М., 1977, с. 40—55.
9. Ипполитов В. Ф. Материалы о межанализаторных различиях по силе нервной системы у человека. — Вопросы психологии, 1966, № 2, с. 23—91.
10. Ипполитов В. Ф. Межанализаторные различия по параметру чувствительности-— силы (возбуждения) для зрения, слуха и кожных ощущений. — В сб.: Типологические особенности высшей нервной деятельности человека. — М., 1967, т. 5, с. 150—167.
118
11. Капустник О. П., Фаддеева В. К. Угашение условных рефлексов у детей от 5 до 15 лет. — В сб.: Опыт систематического исследования условнорефлекторной деятельности ребенка. — М., 1930, с. 19—36.
12. Короткин И. И. Сравнительная характеристика высшей нервной деятельности ребенка при пищевом и оборонительном подкреплении. — В сб.: Основные механизмы условнорефлекторной деятельности ребенка. — М., 1930, с. 95—114.
13. Купалов П. С. Учение о типах высшей нервной деятельности животных. Жури, высш. нервн. деят., 1954, т. 4, вып. 1, с. 3—19.
14. Мерлин В. С. Очерк теории темперамента. — Пермь, 1973.—292 с.
15. Мерлин В. С. Проблемы интегральной характеристики индивидуальности в дифференциальной психофизиологии. — В сб.: Вопросы дифференциальной психофизиологии в связи с генетикой. Пермь, 1974, с. 4—13.
16. Небьшщын В. Д. Индивидуальные различия в зрительном и слуховом анализаторах по параметру сила — чувствительность. — Вопросы психологии, 1957, № 4, с. 53—69.
17. Небылицын В. Д. Электроэнцефалографическое изучение свойства силы нервной системы и уравновешенности нервных процессов у человека с применением факториального анализа. — В сб.: Типологические особенности высшей нервной деятельности человека. М., 1963, т. 3, с. 47—80.
18. Небылицын В. Д. Основные свойства нервной системы человека. — М., 1966. — 384 с.
19. Небылицын В. Д. Психофизиологические исследования индивидуальных различий. — М., 1976. — 336 с.
20. Небылицын В. Д., Голубева Э. А., Равич-Щербо И. В., Ермолаева-Томина Л. Б. Сравнительное изучение кратких методик определения основных свойств нервной системы у человека. — В сб.: Типологические особенности высшей нервной деятельности человека. М., 1965, т. 4, с. 60—84.
21. Павлов И. П. Общие типы высшей нервной деятельности животных и человека.— Избранные труды по физиологии высшей нервной деятельности. М., 1950, с. 199—219.
22. Роговенко Е. С., Соколова Е. В. Угашение с подкреплением как возможный текст для определения уравновешенности нервных процессов у собак. — Журн. высш. нервн. деят., т. 12, вып. 2, с. 279—284.
23. Рождественская В. И., Небылицын В. Д., Борисова М. Н., Ермолаева-Томина Л. Б. Сравнительное изучение различных показателей силы нервной системы у человека. — Вопросы психологии, 1960, № 5, с. 41—56.
24. Русалов В. М. О природе общих и частных свойств нервной системы человека. — В сб.: Психология и психофизиология индивидуальных различий. М., 1977, с. 108—115.
25. Синкевич З. Л. Исследование взаимного внешнего торможения пищевых, оборонительных и исследовательских условных рефлексов. — В сб.: Основные механизмы условнорефлекторной деятельности ребенка. М.; Л., 1930, с. 70—84.
26. Теплов Б. М. Некоторые вопросы изучения общих типов высшей нервной деятельности человека и животных. — В сб.: Типологические особенности высшей нервной деятельности человека. М., 1956, с. 5—123.
27. Федоров В. К. Исследование высшей нервной деятельности собаки безудержного типа. — В сб.: Труды физиологических лабораторий им. И. П Павлова. М.; Л., 1949, т. 15, с. 117—123.
28. Хозак Л. Е. Исследование ориентировочно-исследовательских, пищедобывательных и пассивно-оборонительных условных рефлексов у детей школьного возраста — В сб.: Экспериментальные исследования высшей нервной деятельности ребенка. М., 1933, с. 44—72.
29. Швыркова Н. А., Швырков В. Б. Активность нейронов зрительной коры при пищевом и оборонительном подкреплении. — Нейрофизиология, 1975, т. 7, № 1, с. 100—102.
30. Шугалев Н. П. Соотношение вызванной активности проекционных и ассоциативных зон коры мозга кошки при формировании дифференцировочного торможения пищевого рефлекса. — Журн. высш. нервн. деят., 1971, т. 21, вып. 1, с. 54—59.
31. Strelay I. Problemy i melody baday typow ukladu nerwowego crlewieka. Wrotzlaw—Warszawa—Krakow, 1965. 174 с