149
ЗА РУБЕЖОМ
АДАПТАЦИЯ К ОПТИЧЕСКИМ ТРАНСФОРМАЦИЯМ СЕТЧАТОЧНОГО ИЗОБРАЖЕНИЯ
А. Д. ЛОГВИНЕНКО, Л. Г. ЖЕДУНОВА
Факультет психологии МГУ
Нет нужды доказывать, что проблема научения в восприятии издавна волновала ученых и была одной из центральных в психологии. Достаточно вспомнить, какое место в психологической науке прошлого века занимала дискуссия нативистов и эмпиристов. Идея развития была центральной в системе психологических взглядов Выготского, в теории деятельности Леонтьева, и нам представляется вполне закономерным, что в советской психологии большое внимание уделялось изучению восприятия у детей. Однако изучение онтогенеза восприятия осложняется тем очевидным обстоятельством, что к моменту возникновения у ребенка речи его перцептивные функции мы находим в основном сложившимися и его восприятие уже, по-видимому, ничем не отличается от восприятия взрослого человека. Тем не менее в арсенале методических приемов экспериментальной психологии есть способ, позволяющий взрослому человеку как бы заново пережить процесс перцептивного научения. Эту возможность открывает нам оптическая трансформация сетчаточного изображения. Представляется естественным считать адаптацию к оптической трансформации сетчаточного изображения разновидностью перцептивного научения. Доступность этого процесса самонаблюдению, возможность проведения экспериментов со взрослыми людьми, обладающими развитой рефлексией, делают адаптацию весьма привлекательной моделью для изучения перцептивного научения. За последние двадцать лет адаптация к оптическим искажениям сетчаточного изображения превратилась в широко и глубоко разработанный раздел экспериментальной психологии (где-то на стыке психологии зрительного восприятия и моторного научения) со своими методами и проблематикой. В настоящем обзоре мы попытались отразить наиболее существенные результаты, полученные в этой области, и познакомить советского читателя с современным состоянием этой области экспериментальной психологии, слабо представленной в отечественной психологической литературе.
I. ОСНОВНЫЕ ПОНЯТИЯ И ФАКТЫ
Повторим вслед за Гельмгольцем следующий простой опыт [43]. Пусть на экране перед наблюдателем находится прямая вертикальная линия. Поместив перед глазом наблюдателя клиновидную призму, мы изменим кривизну сетчаточного изображения линии-стимула. Зрительный образ линии, следуя сетчаточному изображению, также будет обладать кривизной. Феномен, однако, заключается в следующем. Если наблюдатель в течение нескольких минут будет пристально смотреть сквозь призму на вертикальную линию, то видимая кривизна линии исчезнет и наблюдатель будет вновь (как и до помещения перед его глазами клиновидной призмы) видеть прямую вертикальную линию. После устранения призмы наблюдателю в течение некоторого времени (1—2 мин) прямая вертикальная линия на экране вновь будет казаться искривленной, но теперь кривизна будет противоположного знака.
Введем некоторые рабочие понятия. Назовем адаптацией к оптическому искривлению линии те процессы, которые совершаются в зрительной системе человека во время наблюдения линии сквозь призму. Термином эффект адаптации будем обозначать результат этих процессов. Иначе говоря, эффект адаптации в описываемом опыте состоит в том, что испытуемый видит прямой искривленную призмой линию. Адаптационным последействием (или просто последействием) будем называть те изменения, которые сохраняются в зрительной системе и после удаления призмы.
Если опыт продолжить и измерить адаптационное последействие по прошествии времени после удаления призмы, мы обнаружим, что последействие уменьшилось, и можно отметить момент времени, когда последействие исчезнет вовсе. Время, в течение которого последействие сохраняется, и те процессы, которые при этом происходят в зрительной системе, будем называть периодом реадаптации и реадаптацией соответственно. В дальнейшем будем различать дистальный и проксимальный стимулы.
150
Дистальный стимул — это реальный физический объект, а проксимальный стимул — это пространственно-временное распределение энергии на сетчатке, вызванное дистальным стимулом.
Мы можем резюмировать теперь итоги описанного опыта: введение закономерной трансформации проксимального стимула посредством оптического устройства (клиновидной призмы) вызывает эффект адаптации, который эквивалентен оптическому преобразованию, но имеет противоположный знак. Адаптационный эффект обладает инерционностью и в отсутствие оптической трансформации требует времени для своего исчезновения.
Оптически вызванная кривизна не является единственной оптической трансформацией, к которой возможна адаптация. В 90-х гг. прошлого века Стрэттон провел ряд исследований, в которых была доказана возможность адаптации к инверсии и реверсии сетчаточного изображения [81]. При инверсии происходит обращение изображения по вертикали, а при реверсии — по горизонтали. Стрэттон с помощью системы линз поворачивал сетчаточное изображение вокруг центра сетчатки на 180° (что создавало разом и инверсию и реверсию изображения). Стрэттон сам был испытуемым и утверждал, что для адаптации к двойной инверсии, т. е. для наступления правильного видения (а именно такого видения, при котором зрительный мир правильно ориентирован относительно гравитационной вертикали), требуется 7—8 дней непрерывного ношения инвертирующих линз (в дальнейшем мы будем говорить: непрерывной экспозиции, к инверсии или вообще к какой-либо оптической трансформации проксимального стимула).
Первоначально интерес психологов к проблеме адаптации сводился к вопросу о возможностях человека адаптироваться к тем или иным оптическим трансформациям. В 1951 г. Колер опубликовал монографию [55], в которой излагались результаты длительных и систематических исследований адаптации к оптическим трансформациям, начатых им еще в 20-е гг. в лаборатории Инсбрукского университета. Эта работа приобрела широкую известность благодаря необычайной изобретательности и остроумию, которые проявил ее автор, демонстрируя поистине неограниченную лабильность зрительной системы при адаптации к различным видам оптической трансформации сетчаточного изображения. Так, Колер в серии хронических экспериментов (т. е. экспериментов с длительной (исчисляемой днями) непрерывной экспозицией к какой-либо оптической трансформации) показал, что адаптация возможна не только к двойной инверсии, но также и к собственно инверсии (без реверсии) и к реверсии (без инверсии). Время, необходимое для полной адаптации как к инверсии, так и к реверсии, составляет 7+2 дня.
Широко распространенным средством трансформации сетчаточного изображения является клиновидная призма. Физически действие клиновидной призмы на луч света состоит в отклонении его траектории на угол, величина которого составляет половину угла при вершине призмы. Однако это простое преобразование сопровождается довольно сложными дополнительными трансформациями сетчаточного изображения. Во-первых, из-за неравномерного отклонения призмой световых лучей различной длины волны вертикальные линии приобретают радужную каемку. Во-вторых, поскольку на величину отклонения луча влияет место его встречи с поверхностью призмы (чем оно ближе к основанию, тем больше линейное смещение луча), возникает нерегулярное искажение контуров и углов.
Колер, будучи испытуемым, носил клиновидные призмы в течение 124 дней. К исходу шестого дня наступила моторная адаптация, т. е. у него полностью восстановилось внешнее поведение (настолько, что он был способен кататься на лыжах). Адаптация перцептивная потребовала более длительного периода. Спустя два дня восстановилась общая стабильность видимого мира (известно, что оптическая трансформация сетчаточного изображения сопровождается утратой константности восприятия в широком смысле этого слова, что переживается испытуемым отчасти и в форме иллюзорного движения зрительного пространства вслед за движением самого испытуемого). Через десять дней исчезли кривизна, искажение углов и радужная каемка. После удаления призмы возникло последействие: было такое впечатление, будто призмы надеты вновь, но их действие имеет обратный знак.
Одним из замечательных достижений Колера было открытие ситуационного последействия [57]. Дело в том, что, когда главная зрительная ось перпендикулярна поверхности призмы, отклонение луча, идущего по главному зрительному направлению, минимально. При смещении глаза величина отклонения луча (а следовательно, и величина искажения) увеличивается. Таким образом, искажение и тем самым сетчаточные изображения предметов зависели от того, куда направлен взор испытуемого. Один и тот же предмет выглядел сжатым, если на него смотреть слева, и растянутым, если смотреть справа. Для уяснения всей сложности такой трансформации следует отметить, что изображение при этом падало на одни и те же участки сетчатки. Механизм адаптации должен, следовательно, основываться на закономерной связи между степенью искажения изображения и положением взора, а не между искажением и местоположением изображения на сетчатке. Ситуационное последействие состоит в том, что по достижении полной перцептивной адаптации и после удаления призм возникали иллюзорные искажения зрительных образов (искривление, растяжение, сжатие и т. п.), величина которых зависела от положения взора. Только зависимость эта носила характер, обратный тому, который имел место в первые часы после надевания призм.
151
В последующих экспериментах Колер еще более усложнил ситуацию. В течение 50 дней испытуемый носил очки, верхняя половина которых представляла собою клиновидные призмы, а нижняя — простые стекла. В зависимости от того, был направлен взгляд — вверх или вниз, испытуемый имел искаженные или неискаженные зрительные образы предметов. Не стоит забывать, что призмы смещают изображение и поэтому, если линия взора проходила сквозь границу, отделяющую верхнюю половину стекла очков от нижней, зрительный образ предмета выглядел разорванным: его верхняя часть была смещена относительно нижней. Оказалось, что адаптация к такой трансформации — дело весьма непростое. Достаточно сказать, что в первые дни появилось последействие при взоре, направленном вниз, т. е. когда испытуемый смотрел на предмет сквозь простое стекло. Однако в дальнейшем это последействие начало уменьшаться и исчезло вовсе. Хотя долгое время после того, как произошла адаптация к верхней части, при взоре, строго направленном на границу между верхней и нижней частью очков, возникало впечатление разорванности образа. После удаления очков последействие такой трансформации держалось длительное время (20—30 дней).
С именем Колера связано открытие еще одного удивительного типа последействий. Выяснилось, что адаптация наступает к оптической трансформации, имеющей место тогда, когда правые половины стекол в очках красные, а левые — зеленые. Если испытуемый смотрит на предмет слева, он видит его зеленым, если справа — красным.
По истечении некоторого времени окраска зрительных образов, вызванная очками, исчезает. После удаления очков возникает цветовое последействие, зависящее от направления взора. При взгляде вправо испытуемый видит мир зеленым, а при взгляде влево — красным. Это явление носит имя эффекта Колера.
Попытки некоторых авторов воспроизвести опыты Колера закончились безрезультатно i[62], другим же удалось не только воспроизвести эффект Колера, но и повторить этот эффект с нейтральными фильтрами вместо цветных [1], [60], 1831. Последействие при этом заключается в том, что после удаления очков при взгляде вправо мир выглядит потускневшим, а влево — просветлевшим.
Оказалось, что не так просто найти такую оптическую трансформацию проксимального стимула, к которой была бы невозможна перцептивная адаптация. Пример такой трансформации мы опять находим у Колера. Речь идет о так называемом цветостереоэффекте. Помещая перед глазами испытуемого клиновидные призмы (основаниями наружу), можно создать своеобразный стереоэффект, если предъявить наблюдателю два монохроматических точечных стимула с различной длиной волны (положим, зеленый и красный точечные источники) и расположенные в одной фронтальной плоскости. Поскольку коротковолновый свет преломляется в большей степени, нежели длинноволновый, монокулярные образы точки будут иметь различные зрительные направления, причем величина диспаратности сетчаточных изображений источника, соответствующего красному цвету, будет противоположного знака сравнительно с диспаратностью сетчаточных изображений монохроматического стимула, соответствующего зеленому цвету. Наличие такой диспаратности и вызывает цветостереоэффект: точечный стимул красного цвета для наблюдателя с призмами будет выглядеть менее, а стимул зеленого цвета — более удаленным относительно плоскости экрана. Вследствие цветостереоэффекта образы вещей пространственно трансформируются: части предмета, имеющие зеленый цвет, будут выглядеть как бы вдавленными в тело предмета, а красные детали объекта, напротив, будут выдавливаться. Один из испытуемых Колера носил клиновидные призмы, создававшие цветостереоэффект, непрерывно в течение 52 дней, и не наблюдалось никаких признаков адаптации.
II. ЭКСПЕРИМЕНТАЛЬНЫЕ ПАРАДИГМЫ
СМЕЩЕННОГО ЗРЕНИЯ
Опыты Колера и его предшественников отличала глобальность подхода. Недостатки этого подхода выявились сразу же. Становилась все более очевидной необходимость сузить постановку вопроса и различать, например, моторную адаптацию и перцептивную. В то же время перцептивная адаптация к различным аспектам одной и той же трансформации протекает по-разному, поэтому имеет смысл различать адаптацию к смещению зрительного направления, которое вызывает клиновидная призма, от искривления контуров, создаваемого той же призмой. Так откристаллизовалось направление, в котором исследуется адаптация лишь к смещению сетчаточного изображения — смещенное зрение. Прежде чем излагать основные результаты в этой области, имеет смысл познакомиться с основными методическими принципами, положенными в основу экспериментального исследования смещенного зрения. Как правило, изучается адаптация к смещению сетчаточного изображения в горизонтальном направлении (латеральное смещение). Существуют три экспериментальные парадигмы смещенного зрения, т. е. три типа экспериментов со смещенным зрением.
1. Экспериментальная парадигма Харриса
Эксперимент в этой парадигме состоит из трех серий [31]. В первой (доэкспозиционной) серии, т. е. до надевания призм, и третьей (постэкспозиционной) серии, т. е. после снятия призм, испытуемому предлагаются три тестовые задачи. Две из них состоят из попадания рукой в цель. Цели бывают двух видов: зрительные и слуховые.
152
В третьей задаче испытуемого просят указать рукой направление «прямо перед собой». В качестве зрительных целей используется ряд вертикальных стерженьков, расположенных во фронтальной плоскости перпендикулярно линии взора испытуемого. Слуховыми целями служат звонки. Во время попадания в слуховую цель и при задаче «прямо перед собой» глаза испытуемого закрыты. При попадании в зрительную цель рука испытуемого двигается под столом так, что он не видит результатов своего попадания.
Основная вторая серия — экспозиционная. Экспозиция к смещенному стимулу длится 3 мин и состоит в следующем. Испытуемому надевают призмы, смещающие его сетчаточное изображение (у Харриса смещение составляло 11 угл. градусов), и он совершает попадания рукой в центральный стержень. При этом рука находится в поле зрения испытуемого, что дает ему возможность видеть результаты своих действий. Мерой адаптации служит разность в точности выполнения тестовых задач в пост- и доэкспозиционной серии.
Таким образом, Харрис в качестве меры адаптации взял послеэффект адаптации (что, кстати, обеспечило возможность сравнения его результатов с фактами, полученными в других экспериментальных парадигмах, в то время как в парадигме Хелда, например, прямое измерение адаптационного эффекта невозможно). В парадигме Харриса адаптация измеряется по трем параметрам. Такими параметрами были: попадание в зрительную цель; в дальнейшем будем называть адаптационный эффект по этому параметру зрительно-моторным адаптационным послеэффектом (ЗМАП); 2) попадание в слуховую цель; 3) определение мануального направления «прямо перед собой»; в дальнейшем будем называть адаптационный эффект по этому параметру проприоцептивным адаптационным послеэффектом (ПАП).
Следует отметить, что при экспозиции (во второй серии) испытуемый совершал движения только одной рукой (некоторые испытуемые — левой, некоторые — правой). Тестовые задачи выполнялись обеими руками. Это позволило ввести еще одну (четвертую) зависимую переменную: интермануальный перенос как разность между величиной адаптационного послеэффекта, взятого по какому-либо из трех параметров, для адаптированной руки и величиной послеэффекта для неадаптированной руки.
В табл. 1 приведены результаты одного из исследований Харриса, выполненного в 1963 г. [31]. Адаптационный эффект по каждому из трех параметров составляет около 50% латерального смещения призм. Это означает, что адаптация была неполной. Интермануальный перенос имеет статистически значимую величину лишь по параметру ЗМАП, хотя и очень незначительную (менее 5%).
В основе теоретической интерпретации Харриса лежит анализ возможной причины адаптационного послеэффекта. Ею не может быть сознательная координация движений руки в момент попадания в цель после того, как испытуемый обнаружил значительную ошибку во время экспозиции к смещению. Ибо в таком случае сознательные коррекции должны прекратиться после удаления призм, и тем самым не должно быть адаптационного послеэффекта. Результаты Харриса не дают возможности считать источником ЗМАП и зрительное смещение, т. е. смещение пространственных координат видимого поля (феноменальное смещение всего видимого поля на величину призматического латерального смещения), поскольку зрительное смещение должно привести к адаптационному послеэффекту на обеих руках, а не только на адаптированной, как это было в эксперименте, о котором идет речь. Не лежит источник адаптивных изменений, по мнению Харриса, и в изменении зрительно-моторных координации, поскольку адаптация не ограничена в данном случае корреляциями между моторным поведением и зрением, а в равной мере затрагивает и слуховую модальность. Величина адаптационного послеэффекта по параметру попадания в слуховую цель лишь немногим меньше ЗМАП (табл. 1).
Адаптационный послеэффект по любому из трех параметров, равно как и практическое отсутствие интермануального переноса, получают свое исчерпывающее объяснение, если предположить наличие проприоцептивного смещения, т. е. изменение чувства положения адаптированной руки. Если говорить совсем просто, при экспозиции к призматическому смещению конфликт между видимым положением руки и ощущаемым (в проприоцептивной модальности) решается в пользу видимого положения: испытуемый начинает чувствовать (проприоцептивно) руку там, где видит. В этом и состоит, по Харрису, суть адаптации к латеральному призматическому смещению сетчаточного изображения. Точка зрения Харриса вошла в литературу по психологии восприятия как гипотеза проприоцептивных изменений.
Важно отметить следующее. Основным феноменом адаптации к призматическому смещению является ЗМАП. Это поведенческая модель смещенного зрения, требующая своей интерпретации. Возможную интерпретацию
Таблица 1
СРЕДНЯЯ (ПО ВОСЬМИ ИСПЫТУЕМЫМ)
РАЗНОСТЬ В ДЮЙМАХ МЕЖДУ ИЗМЕРЕНИЯМИ, СДЕЛАННЫМИ
ДО И ПОСЛЕ АДАПТАЦИИ

* Положительная разность является компенсирующей для призматического смещения (4,6 дюйма). Крестиком помечены статистически значимые отклонения (по Харрису [31]).
153
предложил Харрис: наличие ЗМАП есть результат изменения чувства положения. Изменение чувства положения проявляется в наличии проприоцептивного смешения (ПС), мерой которого служит проприоцептивный адаптационный послеэффект. Итак, гипотеза Харриса может быть кратко сформулирована следующим образом: ЗМАП=ПС, причем ЗС = 0, где ЗМАП — моторный адаптационный послеэффект; ПС — проприоцептивное смещение; ЗС — зрительное смещение.
2. Экспериментальная парадигма Ховарда
Экспериментальная парадигма Ховарда [45], [46] является модификацией парадигмы Харриса. Общая структура эксперимента (до-, пост- и экспозиционные серии) сохраняется. Мерой адаптации также является послеэффект, взятый по какому-либо из трех параметров или по нескольким сразу. Модификации затрагивают экспозицию к призматическому смещению. У Харриса испытуемый во время экспозиции, совершая мануальные движения, имел возможность непрерывно наблюдать за движением своих рук, поэтому такая экспозиция называется экспозицией с полной обратной связью. В парадигме Ховарда испытуемый может видеть лишь кончики своих пальцев в момент попадания в цель, остальная часть руки скрыта от взора испытуемого; рука движется под горизонтальной крышкой стола. Таким образом, испытуемый в порядке обратной связи имеет информацию лишь о конечном результате движения, не имея возможности контролировать его протекание на промежуточных этапах. Поэтому такая экспозиция называется экспозицией с ограниченной обратной связью. Во всем остальном парадигма Ховарда принципиально не отличается от парадигмы Харриса. Ховард, используя эту парадигму, получал 100%-ную адаптацию по параметру ЗМАП (т. е. величина ЗМАП в угловых единицах равнялась угловой величине латерального призматического смещения) [48].
При экспозиции с ограниченной обратной связью помимо проприоцептивного смещения, как правило, получают также и зрительное смещение. Мерой зрительного смещения служит зрительный адаптационный послеэффект (ЗАП). Следует отметить, что тем самым мы вводим еще один параметр адаптации. Операционально зрительный адаптационный послеэффект есть разность в точности выполнения в до- и постэкспозиционной сериях следующей тестовой задачи. В темноте испытуемого просят определить, какая из горизонтального ряда светящихся точек расположена «прямо перед ним». Вариант этой тестовой задачи может состоять в том, что вместо ряда может предъявляться одна движущаяся справа налево (или слева направо) светящаяся точка или пятно, и испытуемый должен вербально отметить, когда стимул будет находиться «прямо перед ним». Ясно, что зрительный адаптационный послеэффект можно трактовать как меру собственно зрительных смещений (ЗС) в результате экспозиции к призматическому смещению.
3. Экспериментальная парадигма Хелда
Своеобразие парадигмы Хелда проистекает из глубокой убежденности автора в том, что адаптация есть проявление более примитивных процессов, чем научение. Такими процессами, по Хелду, являются корреляции между эфферентными и реафферентными сигналами.
Принципиальной особенностью парадигмы Хелда является то, что во время экспозиции к призматическому смещению испытуемый не совершает точностных движений, т. е. движений попадания в цель. Экспозиция состоит в следующем. Испытуемый совершает несколько движений рукой, которая находится в смещенном поле зрения. Движения руки осуществляются на гомогенном фоне, причем поле зрения ограничено, так что испытуемый иногда может даже не догадываться о наличии призматического смещения. До- и постэкспозиционная серии практически такие же, как и в предыдущих парадигмах. Испытуемый совершает попадание в зрительную цель, при этом рука и результаты попадания недоступны испытуемому для наблюдения. Иногда испытуемый просто совершает произвольные перемещения руки в поле зрения. Экспозиция к призматическому смещению длится 3 мин. Существенной особенностью экспериментальной парадигмы Хелда является наличие двух видов экспозиции: активной и пассивной. При активной экспозиции испытуемый совершает активные движения в определенном ритме, который задается щелчками метронома. При пассивной экспозиции совершаются пассивные движения в том же ритме: рука испытуемого перемещается экспериментатором, при этом скорость, траектория и прочие характеристики движения совпадают с характеристиками активных движений.
Основной факт, обнаруженный Хелдом, состоит в том, что адаптация наступает только при активной экспозиции, при пассивной же экспозиции ЗМАП статистически неотличим от нуля. Отметим, что коэффициент адаптации у Хелда значительно ниже, нежели в парадигмах Харриса и Ховарда. В табл. 2 приведены результаты эксперимента Фридмана [24], который сравнил эффективность трех видов экспозиций
Таблица 2
СРЕДНЯЯ ОШИБКА (В ДЮЙМАХ) ПРИ
ПОПАДАНИИ В ЦЕЛЬ ПОСЛЕ АДАПТАЦИИ
К ПРИЗМАТИЧЕСКОМУ СМЕЩЕНИЮ
В 4,08 ДЮЙМА (ПО ФРИДМАНУ [24])
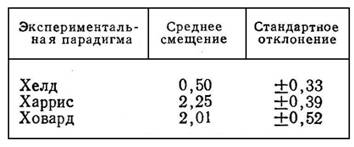
154
(Харриса, Ховарда и Хелда). Правда, и в парадигме Хелда можно получить полную адаптацию. Так, Хелд [37] провел хронический эксперимент со смещенным зрением, в котором в качестве параметра адаптации была взята задача зрительного определения «прямо перед собой». Активная экспозиция состояла в том, что испытуемый с призмами на глазах в естественных условиях самостоятельно передвигался в течение не скольких часов. При пассивной экспозиции испытуемого в принудительном порядке перемещали в той же обстановке, повторяя при этом траекторию и скорость движения «активного» испытуемого. К концу эксперимента все «активные» испытуемые достиг ли 100%-ной адаптации, в противоположность «пассивным» испытуемым, у которых адаптации не было. Поскольку параметром адаптации была успешность зрительного определения направления «прямо перед собой», можно сделать вывод, что в результате адаптации у испытуемых возникло зрительное смещение, равное по величине и обратное по знаку латеральному призматическому смещению.
Теория Хелда, предложенная им для объяснения стабильности зрительного восприятия, позволяет объяснить изменение зрительного восприятия при оптических трансформациях сетчаточного изображения и, кроме того, может служить моделью приобретения индивидуального зрительного опыта. Для изложения его теории требуется ввести новые понятия. Реафферентный сигнал — это некоторое изменение проксимального стимула, причиной которого послужили собственные движения организма. Эксафферентный сигнал — изменение проксимального стимула, вызванное изменением дистального стимула. Таким образом, в общем случае изменение проксимального стимула есть сумма некоторого эксафферент-ного и некоторого реафферентного сигнала. Зрительная система должна уметь расшифровывать этот суммарный сигнал, т. е. вычленять его эксафферентную компоненту. В противном случае наличие реафферентного сигнала будет истолковываться как некоторое изменение дистального стимула и зрение будет источником ложной информации об окружающем мире.
Хелд считает, что вычисление эксафферентной компоненты происходит по такой схеме. Всякий эфферентный сигнал, вызывающий сокращение какой-либо скелетной мышцы, сопровождается побочным центральным разрядом или его эфферентной копией. Эфферентная копия поступает в некоторое гипотетическое устройство, которое Хелд называет коррелятивным накопителем. В коррелятивный накопитель поступает также текущее изменение проксимального стимула (афферентный сигнал), которое параллельно поступает в еще одно гипотетическое устройство — компаратор. В коррелятивном накопителе — своеобразном запоминающем устройстве — происходит отбор следа некоторого афферентного сигнала, а именно того, который наиболее часто в прошлом опыте поступал в коррелятивный накопитель одновременно с данной эфферентной копией. Иначе говоря, актуализируется наиболее коррелирующий с данной эфферентной копией афферентный сигнал. Отобранный в коррелятивном накопителе след афферентного сигнала отсылается в компаратор, где сравнивается с текущим афферентным сигналом. «Исход этого сравнения определяет,— как выражается автор этой модели,— дальнейшие события» [39]. Может, к примеру, происходить вычитание из текущего афферентного сигнала следа того афферентного сигнала, который поступил из коррелятивного накопителя.
Растолкуем теперь, как эта модель может объяснить адаптацию к оптической трансформации. Прежде всего следует отметить, что частота движения глаза гораздо выше, нежели частота смещений объектов, поэтому с данной эфферентной копией в коррелятивном накопителе, как правило, коррелирует соответствующий ей реафферентный сигнал. Таким образом, в компараторе происходит вычитание из афферентного сигнала его реафферентной части, т. е. модель Хелда весьма просто объясняет восстановление эксафферентной компоненты в афферентном сигнале в нормальной ситуации.
Пусть теперь введена оптическая трансформация проксимального стимула, к примеру, призматический сдвиг (α). Это означает, что в отсутствие изменений дистального стимула афферентный сигнал уменьшится на величину (α). Однако коррелятивный накопитель будет отсылать в компаратор старый след, коррелировавший в прошлом опыте с данной эфферентной копией, в результате чего видимое изображение будет смещено на (—α). Однако с течением времени в коррелятивном накопителе все чаще будет подкрепляться новая связь эфферентной копии и все более угашаться старая. В конце концов наступит такой момент, когда данная копия актуализирует новый афферентный след и отошлет его в компаратор для сравнения с текущим афферентным сигналом. Это и будет означать, что произошла адаптация.
Отметим, что основой для адаптации, по Хелду, служит наличие корреляций между эфферентным и реафферентный сигналом; не предполагается никаких связей между абсолютными величинами этих сигналов. В то же время адаптация к любой изменяющейся во времени оптической трансформации проксимального стимула будет практически невозможна, поскольку в этом случае нет основы для корреляции между эфферентным и реафферентный сигналом. Такой вывод из теории Хелда для изменяющихся случайным образом во времени трансформаций проксимального стимула полностью соответствует экспериментальным данным [38].
III. МЕХАНИЗМ АДАПТАЦИИ К СМЕЩЕННОМУ ЗРЕНИЮ
Смещенное зрение вызывает перцептивный конфликт между зрением и другими органами чувств (слухом, осязанием, проприоцепцией
155
и т. д.). Результатом этого интермодального конфликта является разрушение зрительно-моторных координации, проявляющееся в том, что даже такие простые мануальные действия, как попадание рукой в цель, осуществляются с ошибками. Адаптация, т. е. разрешение интермодального конфликта, может происходить либо путем перешифровки сенсорной информации от одной модальности в пользу другой модальности, либо в виде моторного научения. Настоящий раздел посвящен анализу экспериментальных работ, в которых делались попытки выяснить психологический механизм адаптации к смещенному зрению.
Проприоцептивное смещение
Харрис [31], [32], [33] выдвинул гипотезу, согласно которой адаптация к смещению сетчаточного изображения состоит в том, что изменяется воспринимаемое положение кисти руки или какой-либо иной части тела. Тем самым исчезает рассогласование между видимым и проприоцептивным положением руки. Отметим, что, говоря о пространственном чувстве положения, Харрис имел в виду не сенсорно-физиологический уровень, а скорее то, что мы бы назвали перцептивным уровнем. Далее, чувство положения частей собственного тела может обеспечиваться как афферентными сигналами (собственно проприорецептивными сигналами), так и эфферентными (моторными копиями или побочными разрядами) [83]. Тауб и Гольдберг [84] получили значительную адаптацию к призматическому смещению на деафферентированных, т. е. лишенных проприоцепции в узком смысле, обезьянах. Следует все же отметить, что адаптационный послеэффект у деафферентированных обезьян был меньше, чем у нормальных. Это говорит о том, что в изменение чувства положения во время адаптации вносят свой вклад как афферентная, так и эфферентная составляющая.
Позитивным аргументом в пользу существования проприоцептивного смещения руки является наличие проприоцептивного адаптационного послеэффекта (ПАП), который фиксировался в ряде работ [12], [26], [31], [88]. Некоторые авторы, для того чтобы избежать артефактов, связанных с моторным научением, после адаптации просили испытуемых коснуться пальцем неадаптированной руки определенного места на адаптированной руке. В результате испытуемые систематически ошибались [11], [31], [93], [98]. Поскольку в тестовой процедуре Такого рода адаптированная рука вообще не двигалась, ошибки в попадании неадаптированной руки следует отнести целиком за счет изменения чувства положения адаптированной руки. Такую же ошибку обнаружил Харрис [33] при оценке испытуемым расстояния между адаптированной и неадаптированной рукой. Свидетельства в пользу существования проприорецептивного смещения были получены также в задаче попадания рукой в звуковую цель с закрытыми глазами [31], [36], попадания рукой в темноте в большой палец правой ноги [86], а также в задаче установления пассивно (с чужой помощью) перемещающейся руки точно «прямо перед собой» [2] и в задаче установления зрительной цели над воспринимаемым положением невидимой руки [44].
Проприорецептивное смещение возможно между любой парой звеньев в схеме тела, причем необязательно между соседними звеньями. На такую возможность указывали ряд авторов [11], [30, [47], [48]. Крэйск [11] адаптировал испытуемых к призматическому смещению в 20 диоптрий (что соответствует 12 угл. градусам), а затем просил указать неадаптированной рукой плечо, локоть и палец адаптированной руки, скрытой от взора испытуемого. Послеэффект (в сантиметрах) составил для одного испытуемого: плечо — 3,03, локоть — 3,8, палец — 4,6; для другого: плечо — 0,93, локоть — 1,27, палец —2,87. Это означает, во-первых, что в результате адаптации проприорецептивному смещению подверглось не одно звено, а несколько, а во-вторых, более периферийные звенья претерпели более выраженное проприоцептивное смещение. Каханэ и Ауэрбах [51] обнаружили значительно большее проприоцептивное смещение у указательного пальца, который находился в поле зрения во время адаптации, нежели у тех частей тела, которые были не видны во время адаптации. Путтерман с соавторами [72] адаптировал испытуемого, лишив подвижности предплечье, но сохранив ее для кисти. После адаптации он обнаружил изменение чувства положения кисти относительно плеча. Многие авторы считают, что именно воспринимаемое положение руки подвергается перекалибровке при адаптации. Критическим фактом для этой гипотезы является наличие интермануального переноса адаптационного последействия (ЗМАП). Ряд авторов [8], [29], [31], [33], [54] не обнаружили в своих опытах интермануального переноса. Разумеется, наличие интермануального переноса является достаточным условием, чтобы отвергнуть гипотезу изменения чувства положения руки. Однако отсутствие такого переноса еще не является основанием для безоговорочного принятия этой гипотезы [58].
Зрительное смещение
Уже Гельмгольц, который впервые наблюдал адаптацию к призматическому смещению, отмечал наличие переноса адаптационного послеэффекта на неадаптированную руку [43]. Его наблюдения подтверждаются современными исследованиями [9], [10], [52], [77], [88]. Наличие интермануального переноса служит доказательством того, что в результате адаптации к призматическому смещению происходит так называемое зрительное смещение. Выше отмечалось, что критическим тестом для зрительного смещения служит послеэффект в задаче определения зрительного направления
156
«прямо перед собой» (ЗАП), который фиксировался в ряде работ по смещенному зрению [23], [66], [88], [101], [102]. Как смещение зрительного направления можно трактовать наличие послеэффекта в задаче зрительного определения звучащего невидимого объекта [36]. Косвенным свидетельством в пользу существования зрительного смещения можно считать количественное несовпадение зрительно-моторного адаптационного послеэффекта (ЗМАП) и проприорецептивного адаптационного послеэффекта (ПАП) [63], [100].
В основе зрительного смещения может лежать либо изменение локальных знаков сетчатки, т. е. изменение окулоцентрических зрительных направлений относительно сетчаточных координат [8], [47], либо изменение окулоцентрических зрительных направлений относительно эгоцентрического зрительного направления1 [50]. Последнее возможно, если происходит изменение воспринимаемого положения глазного яблока относительно глазницы [17], [52]. Харрис высказал предположение, что во время адаптации изменению может также подвергаться воспринимаемое положение головы [33].
Проприоцептивное смещение положения головы. Колер [56] отмечал, что его испытуемый после длительного ношения смещающих призм, после того как они были удалены, держал голову некоторое время повернутой на 6—9 угл. градусов в направлении призматического смещения, не осознавая этого. Аналогичное наблюдение сделал и Харрис [31], [33]. Объективным Доказательством существования проприоцептивного смещения положения головы после адаптации к смещению сетчаточных изображений считают наличие послеэффекта в задаче «выставить прямо перед собой» источник звука в темноте. Наличие такого послеэффекта было показано в ряде работ [22], [25], [75].
Проприоцептивное смещение положения глаза. Некоторым авторам удалось зарегистрировать положение глаза при инструкции «смотреть прямо перед собой» до адаптации, во время адаптации и после адаптации. Были обнаружены существенные различия между положением глаза до адаптации и во время адаптации [64], а также после адаптации [13], [53]. Хотя положение головы при этом не регистрировалось, однако аналогичные результаты были получены в условиях, когда во время адаптации движения головы ограничивались, что делало маловероятным проприоцептивное смещение головы [4], [62].
Косвенным подтверждением существования такого явления, как изменение воспринимаемого положения глаза, является наблюдение Колера [55], сделанное им после того, как у испытуемого были удалены очки, смещавшие верхнюю половину поля зрения относительно нижней. По просьбе совершить движение глазами строго вертикально сверху вниз этот испытуемый совершал зигзагообразные движения. Иначе говоря, траектория движения глаза при этом состояла из двух вертикальных отрезков, смещенных друг относительно друга в горизонтальном направлении. Такие же данные с помощью фотографирования получили Пик и другие [69].
Аддитивная модель
В своем теоретическом анализе возможных механизмов адаптации к смещению Харрис [33] высказал предположение о том, что зрительно-моторный адаптационный по-слеэффект есть алгебраическая сумма про-приорецептивного и зрительного смещения, Ранее эту же мысль высказывал Гамильтон [29]. Уилкинсон [102] сформулировал это утверждение в виде так называемой линейной аддитивной модели: ЗМАП = ПС + ЗС, где ЗМАП — зрительно-моторный послеэф-фект; ПС — проприорецептивное смещение, ЗС — зрительное смещение. Экспериментальные данные некоторых авторов [36], [63], [64], [73], [74], [90], [91] и собственные данные Уилкинсона подтверждают эту модель. Уэлч [99] предложил трехкомпонент-ную линейную модель адаптационного послеэффекта. В его экспериментах [101], а также в экспериментах некоторых других авторов [2], [86], [92] сумма зрительного и проприорецептивного адаптационных после-эффектов (ЗАП-Ь ПАП) составляла несколько меньшую величину, нежели зрительно-моторный адаптационный послеэф-фект. Остаток (третья компонента) указывает на некий третий источник адаптационных изменений, природа которого, на наш взгляд, в полной мере не ясна. По мнению Уэлча, он связан с моторным научением во время адаптации.
Адаптация как моторное научение
Уже при появлении самых первых экспериментальных работ по адаптации к оптическим искажениям некоторые авторы, находившиеся в ту пору под влиянием бихевиоризма, высказывали взгляд на адаптацию как разновидность моторного научения [18], [19], [20], [80], [103]. Исторический анализ работ по адаптации к оптическим искажениям с этих позиций дали Смит и Смит [78]. Адаптация трактуется ими как формирование новых комбинаций моторных команд. Наличие изменений в сенсорной плоскости этими авторами либо полностью отрицается [78], либо объявляется иллюзиями [18], либо результатами «переименования» [21], [28].
В настоящее время трудно сомневаться в том, что в ситуации экспериментальной парадигмы Харриса (так же как и Ховарда) имеет место некоторое моторное научение. Тауб даже придерживается той точки зрения, что адаптационный эффект может быть полностью истолкован как результат
157
оперантного научения, а именно избирательного оперантного научения [83]. Его позиция состоит в том, что интермодальный конфликт между зрительной и проприоцептивной информацией сам по себе может служить негативным подкреплением второго рода для реакции избегания, поскольку в прошлом опыте (индивидуальном, равно как и видовом) такого рода конфликты, как правило, сопровождались сильными биологически отрицательными событиями (подкреплениями первого рода). Поэтому в соответствии с законами оперантного обучения и накапливаются все реакции, уменьшающие величину интермодального конфликта. Имеется солидное экспериментальное обоснование такой точки зрения. Тауб и Гольдберг [84] провели изящный эксперимент, направленный непосредственно на доказательство гипотезы об адаптации как виде избирательного оперантного научения. Испытуемому надевали призматические очки, рука его помещалась на подлокотнике, и испытуемый совершал движения рукой справа налево, слева направо, назад — вперед и наоборот. При этом рука испытуемого постоянно находилась в поле его зрения. Величина латерального смещения призм могла изменяться в зависимости от скорости перемещения руки испытуемого (вместе с подлокотником). Изменение величины призматического смещения происходило в паузах между сериями движений испытуемого. Серия состояла из упомянутых выше четырех движений. Если испытуемый увеличивал скорость движения от серии к серии, величина призматического смещения уменьшалась, если скорость движения не изменялась или уменьшалась, величина призматического смещения увеличивалась. Испытуемые, которые даже и не подозревали о том, что их поле зрения смещено, и которым давалась совершенно нейтральная инструкция перемещать руку, без всякого указания на скорость движения все без исключения постепенно уменьшали призматическое смещение до нуля. Более того, этот результат не зависел от того, в каком отношении изменение призматического смещения находилось к скорости движения: аналогичный результат получался, если призматическое смещение увеличивалось при увеличении скорости.
Тот факт, что все реакции, даже самые незначительные по абсолютной величине, удерживаются и закрепляются, если они уменьшают интермодальный конфликт, находит свое подтверждение в методике «призматического оформления», предложенной Ховардом [45]. Суть этой методики состоит в том, что призматическое смещение экспонируется испытуемому не внезапно, а постепенно и равномерно. Начиная с нулевого смещения через равные промежутки времени призматическое смещение увеличивается на некоторую величину до тех пор, пока не достигнет заданного уровня (10°—20°). Замечательная особенность этой методики состоит в том, что шаг изменения (приращение) призматического смещения не превышает величину средней ошибки попадания в цель испытуемым в нормальных условиях, т. е. испытуемый не имеет возможности сознательно обнаружить наличие призматического смещения. Тем не менее в этих условиях происходит адаптация, равно как и в ситуации классической парадигмы Ховарда. Более того, коэффициент адаптации (отношение величины адаптационного эффекта к величине призматического смещения) при «призматическом оформлении» также равен 100%.
Уэлч [98] представил сводку данных в пользу того, что адаптация есть научение. Во-первых, он проследил общие черты, которыми обладают адаптация и моторное научение, во-вторых, он привлек данные других авторов и показал, что так называемые классические факторы, влияющие на научение, оказывают влияние и на адаптацию и, в-третьих, он показал, что мотивация и подкрепление оказывают такое же влияние на адаптацию, как и на научение.
Факторы, определяющие вид и место адаптивных смещений
Выше было показано, то адаптивные смещения можно различать по виду (зрительные и проприоцептивные) и по месту (в каком звене сенсорной плоскости они произошли). Адаптивные смещения различного вида могут одновременно происходить в различных местах. Возникает вопрос о факторах, которые определяют, какого вида и в каком месте произойдет адаптивное смещение в данных экспериментальных условиях. Большинство авторов, которым не удалось получить интермануальный перенос, использовали экспериментальную парадигму Харриса [8], [9], [10], [11], [29], [31], [33], [54], [65], [97]. И наоборот, наличие интермануального переноса было получено в экспериментальной парадигме Ховарда [2], [5], [24], [27], [45], [50], [52], [94], [95], [96]. Напомним, что в парадигме Харриса испытуемый имеет возможность наблюдать смещение своей руки в течение всего адаптационного периода, в то время как в парадигме Ховарда испытуемый видит лишь кончики своих пальцев в конечной фазе действия.
Некоторые авторы сравнивали специально эти условия адаптации и пришли к выводу, что для наличия интермануального переноса необходимо, чтобы испытуемый не видел свою руку смещенной во время адаптации [9], [80], [88]. Следовательно, если испытуемый видит смещенной руку во время адаптации, последняя завершается проприоцептивным смещением, если не видит — зрительным смещением. Этот вывод подтверждается также следующими фактами. Хронические эксперименты с призматическим смещением, в которых испытуемый свободно перемещался в привычном окружении и мог, естественно, без ограничений наблюдать все части своего тела, попадавшие в поле зрения, завершались 100%-ным зрительным смещением [36], [77]. Зрительное
158
смещение фиксировали в экспериментах, где во время адаптации испытуемый мог наблюдать свои неподвижные ступни или туловище [14], [15], [16] или свои ноги в движении [65], [66].
Кэнон [6], [7] высказал гипотезу, которую развивал далее совместно с Юлариком [88], что адаптивное смещение возникает в той модальности, которой испытуемый уделяет меньше внимания. Аналогичное предположение делал ранее Кохен [9].
Фридман и Рэкош [25] провели интересный эксперимент, который проливает свет на вопрос относительного места, где происходят адаптивные изменения. Их испытуемые носили одну призму, смещавшую зрительное направление на 10 угл. градусов, и во время адаптационного периода, длившегося 4 ч, свободно перемещались. У половины испытуемых на ушах были наушники. Во время адаптации каждые полчаса тестировались зрительное и слуховое направления «прямо перед собой». Те испытуемые, у которых были наушники, показали преимущественное проприоцептивное смещение головы, в то время как вторая группа испытуемых (без наушников) показала проприоцептивное смещение глаза. Вероятно, адаптивное изменение головы более предпочтительно тогда, когда нет слуховых стимулов и нет поэтому конфликтов, состоящих в том, что объект видится не там, где слышится.
В контексте обсуждения условий, которые вызывают те или иные адаптивные смещения, интерес представляют следующие феномены. Во-первых, Рок с соавторами [76] сообщали о возможности получить смещение зрительного эгоцентрического направления во время очень кратковременного рассматривания окружения через смещающую призму без каких бы то ни было перемещений тела и его конечностей, которые к тому же оставались за пределами поля зрения. Эту «мгновенную адаптацию» к смещению можно было зафиксировать в виде зрительного адаптационного послеэффекта. Во-вторых, часто испытуемые вовсе не замечают рассогласования между зрением и чувством положения руки в таких ситуациях. Они просто ощущают руку там, где видят. Это явление получило название «зрительная ловушка» [35], [85] и детально исследовалось [69], [70], [96]. Явление зрительной ловушки часто приводится в качестве доказательства изначального доминирования зрения над осязанием. Зрительная ловушка наблюдается при небольших (до 10 угл. градусов) смещениях и быстро исчезает при отключении зрения. Аналогичное доминирование зрения над слухом отмечали Ховард и Тэмплтон [49]. В-третьих, Харрис [34] заметил, что возможно «когнитивное смещение» эгоцентрического зрительного направления и для этого не требуется рассогласования между сенсорной информацией в зрительной и других модальностях. По мнению Харриса, «мгновенная адаптация», которую наблюдал Рок, связана с когнитивным смещением.
Необходимость активных движений для адаптации к
смещению зрительного направления
Теория адаптации Хелда предполагает установление новых корреляций между моторными копиями (или побочными разрядами) и реафферентными сигналами. Для этого необходимы активные движения, но совсем необязательно осознание конфликта между зрением и проприорецепцией. Последнее обстоятельство было убедительно доказано в эксперименте с адаптацией к кривизне, где адаптационный послеэффект был получен после пребывания испытуемого в помещении, стены которого оклеены бумагой со случайно-точечными узорами. Такой характер зрительного окружения оставлял испытуемого в полном неведении относительно того, что его зрительное направление было искажено [41].
Вывод о необходимости активных движений сделан Хелдом на основании результатов его многочисленных работ, где было показано, что адаптационный послеэффект возникает лишь при наличии активных движений в поле зрения, характер действия при этом не важен. Разумеется, эти опыты были проведены в экспериментальной парадигме Хелда. Адаптационный послеэффект возникал, если испытуемый в течение двух минут адаптационного времени произвольно перемещал свою руку в поле зрения [40] или если адаптация состояла в том, что испытуемый должен был написать свое имя в смещенном поле зрения [37], [38], [40], [42].
Однако ряд исследователей показал все же, что адаптации можно достичь без активных движений [23], [68], [79]. Эти авторы считают, что значительное различие между активными и пассивными условиями в опытах Хелда с соавторами вызвано различием между экспериментальными условиями, в которых проходила адаптация и тестирование послеадаптационных эффектов.
Другая точка зрения состоит в том, что в парадигме Хелда при пассивной адаптации у испытуемого меньше информации, чем при активной адаптации. Если при пассивной адаптации испытуемому достаточно информации о смещении, можно добиться того же эффекта, что и при активной адаптации [13], [59].
1. Abramovitz A. A computer-controlled conditioning procedure designed to produce gaze-contingent shifts in color perception. — S. A. J. Psychol., 1973, v. 3, p. 47—72.
2. Baily J. S. Adaptation to prisms: Do proprioceptive changes mediate adapted behavior with ballistic arm movoments? — Quart. J. Exp. Psychol., 1972, v. 24, p. 8—20.
3. Baily I. S. Arm-body adaptation with passive arm movements. — Percep. and Psychophys., 1972, v. 12, p. 39—44.
4. Bossom J. Mechanism of prism adaptation in normal monkeys. — Psychonomic Science, 1964, v. 1, p. 377—378.
159
5. Bossom J., Hamilton С R. Interocular transfer of prism-altered coordinations in split-brain monkeys. — J. Сотр. and Physiol. Psychol., 1963, v. 56, p. 769—774.
6. Canon L. K. Intermodality inconsistency of input and directed attention as determinants of the nature of adaptation. — J. Exp. Psychol., 1970, v. 84, p. 141—147.
7. Canon L. K- Directed attention and maladaptive adaptation to displacement of the visual field. — J. Exp. Psychol., 1971, v. 88, p. 403—408.
8. Cohen H. B. Some critical factors in prism adaptation. — Amer. J. Psychol., 1966, v. 79, p. 285—290.
9. Cohen M. M. Continuous versus terminal visual feedback in prism aftereffects. — Percept, and Mot. Skills, 1967, v. 24, p. 295— 1302.
10. Cohen M. M. Changes in auditory localization following prismatic exposure under continuous and terminal visual feedback. — Percept, and Mot. Skills, 1974, v. 38, p. 1202.
11. Craske B. Change in transfer function of joint receptor output. — Nature, 1966, v. 210, p. 704.
12. Craske B. Intermodal transfer of adaptation to displacement. — Nature, 1966, v. 210, p. 765.
13. Craske B. Adaptation to prisms-change in internally registered eye position. — Brit. J. Psychol., 1967, v. 58, p. 329—330.
14. Craske В., Crawshaw M. Adaptive changes of opposite sign in the oculomotor system of the two eyes. — Quart. J. Exp. Psychol., 1974, v. 26, p. 106—113.
15. Craske В., Templeton W. B. Prolonged oscillation of the eyes induced by conflicting position input. — J. Exp. Psychol., 1968, v. 76, p. 387—393.
16. Crawshaw W., Craske B. No retinal component in prism adaptation. — Acta Psychologica, 1974, v. 38, p. 421—423.
17. Ebenholtz S. M. The possible role of eye-muscle potentiation in several forms of prism adaptation. — Perception, 1974, v. 3, p. 477—485.
18. Ewert P. H. A study of the effect of inverted retinal stimulation upon spatially coordinaled behavior. — Genetic Psychol. Monographs, 1930, v. 7, p. 177—363.
19. Ewert P. H. Factors in space localization during inverted vision. I. Interference. — Psychol. Rev., 1936, v. 43, p. 522—546.
20. Ewert P. H. Factors in space localization during inverted vision. 2. An explanation of interference and adaptation. — Psychol. Rev., 1937, v. 44, p. 105—106.
21. Festinger L. Burnham С A., Ono H., Bamber D. Efference and the conscious experience of perception. — J. Exp. Psychol. Monograph., 1967. v. 74(4), p. 1—36.
22. Fishkin L. Head adaptation to lateral displacement. — Percept, and Mot. Skills, 1969, v. 29, p. 291—297.
23. Foley J. E., Maynes F. J. Comparison of training methods in the production of prism adaptation.—J. Exp. Psychol., 1969, v. 81, p. 151—155.
24. Freedman S. J. Perceptual compensation and learning. — In: Freedman S. J. (ed.) The neuropsychology of spatially oriented behavior. Homewood: Dorsey Press, 1968, p. 63—76.
25. Freedman S. J., Rekosh J. H. The functional integrity of spatial behavior. In: Freedman S. J. (ed.). The neuropsychology of spatially oriented behavior. Homewood: Dorsey Press, 1968, p. 153—162.
26. Freedman S. I., Wilson L. Compensation for auditory rearrangement. — Percept, and Mot. Skills, 1967, v. 26, p. 861—866.
27. Goldberg I. A., Taub E., Berman A. J. Decay of prism aftereffect and interlimb transfer of adaptation. Paper presented at the Easter Psychological Association. Boston, 1967.
28. Gyr I. W., Wdley R. The effect of efference to the arm on visual adaptation to curvature: A replication. — Psychon. ScL, 1970, v. 21, p. 89—91.
29. Hamilton C. R. Intermanual transfer of adaptation to prisms. — Am. J. Psychol., 1964, v. 77, p. 457—462.
30. Hamilton С R., Sullivan M. V., Hillyard S. A. Effects of adaptation to displaced vision on reaching to remembered position (unpublished). Standford University, Stand-ford/University of California, San Diego, 1971.
31. Harris C. S. Adaptation to displaced vision: Visual, motor, or proprioceptive change? — Science, 1963, v. 140, p. 812—813.
32. Harris C. S. Comment of "change in straight ahead". — Psychon. Sci., 1965, v. 2, p. 285.
33. Harris С S. Perceptual adaptation to inverted, reversed and displaced vision. — Psychol. Rev., 1965, v. 72, p. 419—444.
34. Harris С S. Beware of the straight-ahead shift, a nonperceptual change in experiments on adaptation to displaced vision. — Perception, 1974, v. 3, p. 461—476.
35. Hay J. C, Pick H. L., Jr., Ikeda K-Visual capture produced by prism spectacles. — Psychon. Sci., 1965, v. 2, p. 215—216.
36. Hay J. C., Pick H. L., Jr. Visual and proprioceptive adaptation to optical displacement of the visual stimulus. — J. Exp. Psychol., 1966, v. 71, p. 150—158.
37. Held R. Exposure history as a factor in maintaining stability of perception and coordination. — J. Nerv. Ment. Dis., 1961, v. 132, p. 26—32.
38. Held R., Freedman S. Plasticity in human sensorimotor control. — Science, 1963, v. 142, p. 455—461.
39. Held R., Hein A. A neural model for labile sensorimotor coordination. — In: Bernard E., Hare M. (eds.) Biological prototypes and synthetic systems. V. I. N. Y.: Plenum Press, 1962, p. 71—74.
40. Held P., Gottlieb N. Adaptation of disarranged hand-eye coordination. — Percept, and Мог. Skills, 1958, v. 8, p. 83—86.
41. Held R., Rekoch J. Motor-sensory feedback and the geometry of visual space. — Science, 1963, v. 141, p. 722—723.
42. Held R., Schlank M. Disarranged eye-hand coordination. — Am. J. Psychol., 1959, v. 72, p. 602—605.
43. Helmholtz H. von. Handbuch der Physiologishen
160
Optik. Leipzig: Voss, 1867. 874 S.
44. Hillyard S. A., Hamilton С R. Misloca-lization of the arm following adaptation to displaced vision (unpublished). University of California, San Diego/Stanford University, Stanford, 1971.
45. Howard I. P. Response shaping to visual-motor discordance. Paper presented to the Psychonomic Society, Chicago, 1967.
46. Howard I. P. Displacing the optical array. — In: Freedman S. J. (ed.) The neuropsychology of spatially oriented behavior. Homewood: Dorsey Press, 1968, p. 19—36.
47. Howard I. P. The adaptability of the visual-motor system. — In: Connolly К. Н. (ed.) Mechanisms of motor skill development. London: Academic Pres, 1970, p. 337—352.
48. Howard I. P. Perceptual learning and adaptation. — Brit. Med. Bull., 1971, v. 27, p. 248—252.
49. Howard I. P., Tetnpleton W. B. Human spatial orientation. N. Y.: John Wiley, 1966.
50. Howard I. P., Anstis Т., Lucia H. C. The relative lability of mobile and stationary components in a visual-motor adaptation task. — Quart. J. Exp. Phychol., 1974, v. 26, p. 293—300.
51. Kahane J., Auerbach C. Effect of prior body experience on adaptation to visual displacement. — Perception and Psychophysics, 1973, v. 13, p. 461—466.
52. Kalil R. E., Freedman S. J. Intermanual transfer of compensation for displaced vision.—Percept, and Mot. Skills, 1966 a, v. 22, p. 123—126.
53. Kalil R. E., Freedman S. J. Persistence of ocular rotation following compensation for displaced vision. — Percept, and Mot. Skills, 1966b, v. 22, p. 135—139.
54. Kennedy J. Prismatic displacement and the remembered location of targets. — Perception and Psychophysics, 1966, v. 5, p. 218— 220.
55. Kohler I. Uber Aufbau und Wandlungen der Wahrnehmungswelt, insbesondere fiber "bedingte" Empfindungen. Rohrer, Vienna, 1951. 118 S.
56. Kohler I. Experiments with prolonged optical distortions. — Acta Psychologica, 1955, v. 11, p. 176—178.
57. Kohler I. Experiments with goggles. — Scient. Amer., 1962, v. 206, p. 62—86.
58. Kornheiser A. S. Adaptation to laterally displaced vision: A review. — Psychol. Bull., 1976, v. 83, p. 783—816.
59. Kravitz I. H., Wallach H. Adaptation to displaced vision contingent upon vibrating stimulation. — Psychon. Sci., 1966, v. 6, p. 465—466.
60. Leppmann P. K., Wieland B. A. Visual distortion with two-colored spectacles. — Percept, and Mot. Skills, 1966, v. 23, p. 1043— 1048.
61. McCollough C. The conditioning of color-perception. — Amer. J. Psychol., 1965, v. 78, p. 362—378.
62. McLaughlin S., Bower J. Auditory localization and judgements of straight ahead during adaptation to prisms. — Psychon. Sci., 1965, v. 2, p. 283—284.
63. McLaughlin S. C, Rifkin K. L., Webster R. G. Oculomotor adaptation to wedge prisms with no part of the body seen. — Perception and Psychophysics, 1966, v. 1, p. 452—458.
64. McLaughlin S. C., Webster R. G. Change in straight-ahead eye position during adaptation to wedge prisms. — Perception and Psychophysics, 1967, v. 2, p. 37—44.
65. Mikaelian H. H. Adaptation to rearranged eye-foot coordination. Paper presented at the Eastern Psychological Association, N. Y., 1966.
66. Mikaelian H. H. Adaptation to rearranged eye-foot coordination. — Perception and Psychophysics, 1970, v. 8, p. 222—224.
67. Mikaelian H. H. Gsneralized sensorimotor adaptation with diminished feedback. — Psychologische Forschung, 1974, v. 36, p. 321—328.
68. Pick H. L., Jr., Hay J. С. A passive test of the Held reafference hypothesis. — Percept, and Mot. Skills, 1965, p. 20, p. 1070—1072.
69. Pick H. L., Jr., Hay J. C, Martin R. Adaptation to splitfield wedge prism spectacles. — J. Exp. Psychol., 1969, p. 80, v. 125— 132.
70. Pick H. L., Warren D. H., Mclntyre C., Appel L. Transfer and the organization of perceptual-motor space. — Psychologishe Forschung, 1972, v. 35, v. 163—177.
71. Polyak S. The vertebrate visual system. Chicago: University of Chicago Press, 1957. 1390 p.
72. Putterman A. H., Robert A. L., Bregman A. S. Adaptation of the wrist to displacing prisms. — Psychon, Sci., 1969, v. 16, p. 79—80.
73. Redding G. M. Additivity in adaptation to optical tilt. — J. Exp. Psychol: Human Perception and Performance, 1978, v. 4, p. 178—190.
74. Redding G. M., Wallace B. Components of displacement adaptation in acquisition and decay as a function of hand and hall exposure. — Perception and Psychophysics, 1976, v. 20, p. 453—459.
75. Rekosh J., Freedman S. J. Errors in auditory direction-finding after compensation for visual rearrangement. — Perception and Psychophysics, 1967, v. 2, p. 466—468.
76. Rock I., Goldberg J., Mack .A. Immediate correction and adaptation based on viewing a prismatically displaced scene, — Perception and Psychophysics, 1966, v. 1, p. 351—354.
77. Shaffer O., Wallach H. Adaptation to prismatic displacements: Hand position and target location. — J. Exp. Psychol., 1966, v. 72, p. 207—212.
78. Smith K- U., Smith W. K. Perception and motion. Philadelphia: Saunders, 1962. 341 p.
79. Singer G., Day R. Spatial adaptation and aftereffect with optically transformed vision effects of active and passive responding and the relationship between test and exposure responses. — J. Exp. Psychol., 1966, v. 71, p. 725—731.
80. Snyder F. W., Pronko N. H. Vision with spatial inversion. Wichita: University of
161
Wichita Press, 1952. 144 p.
81. Stratton G. Upright vision and the retinal image. — Psychol. Rev., 1896, v. 4, p. 182—187.
82. Stubbs A., Abramovitz A. Adaptation to split-field dual-density achromatic spectacles and its perceptual aftereffect. — S. A. J. Psychol., 1974, v. 4, p. 10—15.
83. Taub E. Prism compensation as a learning phenomenon: A philogenetic perspective. — In: Freedman S. J. (ed.). The neuropsychology of spatially oriented behavior. Homewood: Dorsey Press, 1968, p. 77—106.
84. Taub E., Goldberg I. A. Use of sensory recombination and somatosensory deafferentation techniques in the investigation of sensory-motor integration. — Perception, 1974, v. 3, p. 393—408.
85. Tastevin J. En partant de l'experience d'Aristote. — L'Encephale, 1937, v. 1, p. 57—158.
86. Templeton W. В., Howard I. P., Wilkinson D. A. Additivity of components of prismatic adaptation. — Perception and Psychophysics, 1974, v. 15, p. 249—257.
87. Uhlarik J. J. Role of cognitive factors on adaptation to prismatic displacement. — J. Exp. Psychol., 1973, v. 98, p. 223—232.
88. Uhlarik J. J., Canon L. K. Influence of concurrent and terminal exposure conditions on the nature of perceptual adaptation. — J. Exp. Psychol., 1971, v: 91, p. 233—239.
89. Wallach H., Moore M. E., Davidson L. Modification of stereoscopic depth perception. — Amer. J. Psychol., 1963, v. 76, p. 191—204.
90. Wallace B. Stability of Wilkinson's linear model of prism adaptation over time for various targets. — Perception, 1977, v. 6, p. 145—151.
91. Wallace В., Fisher L. E. Proprioception and the production of adaptation and interma-nual transfer to prismatic displacement. — Perception and Psychophysics, 1979, v. 26, p. 113—117.
92. Wallace В., Redding G. M. Additivity in prism adaptation as manifested in inter-manual and interocular transfer. — Perception and Psychophysics., 1979, v. 25, p. 133—136.
93. Wallach H., Huntington D. Counteradaptation after exposure to displaced vision direction. — Perception and Psychophysics, 1973, v. 13, p. 519—524.
94. Walsh T. M., Freedman S. J. Ipsilateral hand-eye rearrangement and controlateral transfer compared under differential adaptation criteria. Paper presented at the Eastern Psychological Association. N. Y., 1966.
95. Walsh T. M., Freedman S. J. Intermanual transfer of compensation for prism displacement in the absence of error-corrective feedback. Paper presented at the Eastern Psychological Association. Boston, 1967.
96. Warren D. H., Cleaves W. T. Visual-proprioceptive interaction under large amounts of conflict. — J. Exp. Psychol., 1971, v. 90, p. 206—214.
97. Warren D. H., Piatt В. В. The subject: A neglected factor in recombination research. — Perception, 1974, v. 3, p. 421—438.
98. Welch R. B. Adaptation to prism-displaced, vision: The importance of target pointing. — Perception and Psychophysics, 1969, v. 5, p. 305—309.
99. Welch R. B. Speculation on a model of prism adaptation. — Perception, 1974, v. 3, p. 367—392.
100. Welch R. В., Rhoades R. IT..The manipulation of Informational feedback and its effects upon prism adaptation. — Canad. J. Psychol., 1969, v. 23, p. 415—428.
101. Welch R. В., Choe С S., HeinrichD. R, Evidence for a three-component model of prism adaptation. — J. Exp. Psychol., 1974, v. 103, p. 700—705.
102. Wilkinson D. A. Visual-motor control loop: A linear system? — J. Exp. Psychol., 1971, v. 89, p. 250—257.
103. Wooster M. Central factors in the development of a new spatial coordination. — Psychol. Monogr., 1923, v. 32(4).
1 Эгоцентрическое зрительное направление определено в полярной системе координат, центр которой связан с головой (а не с сетчаткой, как это имеет место для окулоцентрического зрительного направления).