116
КОРОТКИЕ СООБЩЕНИЯ
НЕЙРОФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ИДЕОМОТОРНОЙ ТРЕНИРОВКИ
Л. ПИККЕНХАЙН
Лейпциг
Под идеомоторной тренировкой мы понимаем повторяющийся процесс интенсивного представления движения, воспринимаемый как собственное движение, который может способствовать выработке, стабилизации и исправлению навыков и ускорить их развитие в практической тренировке.
Сегодня виды идеомоторной тренировки широко применяются — сознательно или бессознательно — в физиологии труда, космической физиологии, физиологии спорта, физиологии системы человек — машина, в других областях прикладной физиологии. Их применение важно особенно там, где временно нельзя осуществить действительное выполнение, т. е. реальную тренировку, соответствующих сложных движений.
Уже давно известно, что интенсивное мышление о движении способствует тенденции его выполнения. Идеомоториый феномен — появление биотоков мышц во время интенсивного воображения выполнения действий — был описан уже в начале этого столетия. В своей статье «Физиологический механизм так называемых произвольных движений» И. П. Павлов [4] писал в 1936 г.: «Давно было замечено и научно доказано, что, раз вы думаете об определенном движении (т. е. имеете кинэстезическое представление), вы его невольно, этого не замечая, производите».
Не удалось найти никаких литературных данных о нейрофизиологических механизмах, на которых основываются идеомоториый феномен как частичный и идеомоторная тренировка как общий процесс. Каждое действие организма требует предварительного программирования движений, в основе которого лежит формулировка цели действия. Основные работы по этому вопросу были сделаны Н. А. Бернштейном и П. К. Анохиным. Реализация этой программы действия осуществляется через подпрограммы различной сложности, иннервационные паттерны которых складываются иерархически на различных уровнях центральной нервной системы. В зависимости от задачи, которая должна быть разрешена, эти иерархически расчлененные иннервационные паттерны употребляются в рамках филогенетически запечатленных функциональных систем [1], [2].
По принципам функциональной системы, разработанным П. К. Анохиным, они по мере надобности включают члены обратной связи, которые обеспечивают беспрерывный контроль и динамическое приспосабливание элементов действия к данной ситуации, чтобы достигнуть в программе действия заданную конечную цель. При каждой тренировке двигательных навыков обратные связи, сигнализирующие конечный результат движения, играют существенную роль. При этом беспрерывно сопоставляются реафферентные информации с эфферентной копией [7]. В качестве реафферентных сигналов о результате реального движения действуют при этом сложные информации от экстерорецепторов, особенно от оптического, акустического, тактильного и вестибулярного анализаторов, а также от проприорецепторов в суставах, сухожилиях и мышцах.
Решающими основными факторами для выполнения действия в рамках функциональной системы, а также для двигательного обучения являются следующие:
1.Управляющая двигательная программа или программа действия, которая у человека может быть в различной степени осознана.
2.Программой действия определяются двигательные подпрограммы или центральные иннервационные паттерны различной степени интеграции.
3.Обратная сигнализация о результате действия, сопоставляемая с эфферентной командой программы действия и подводящая нужную корректуру.
Если рассмотреть идеомоторное воспроизведение двигательного навыка, т. е. его воображаемое выполнение в воображаемом пространстве действия, то факторы 1 и 2 существуют, а фактор 3 — обратная информация о выполнении движений — отсутствует. Как в этих условиях можно объяснить нейрофизиологические механизмы неоспоримых эффектов идеомоторной тренировки?
Экспериментальные морфологические и нейрофизиологические исследования последних лет показали, что кроме обратных связей экстеро- и проприорецепторов, которые мы хотим назвать внешними обратными связями, существуют еще внутренние обратные связи, осуществляемые различными уровнями организации иерархической архитектуры
117
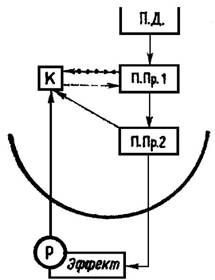
Рис. 1. «Структурная единица» внутренних обратных связей эфферентной моторной системы изображена на примере уровня подпрограммы 1-го и 2-го порядка. П. Д. — программа действия; ППр. 1 в ППр 2 — подпрограммы действия 1-го и 2-го порядка; Р — рецептор; К — копия эфферентного иннервационного паттерна.
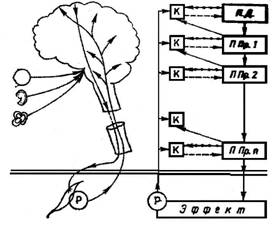
Рис. 2. Схематическое изображение внутренних обратных связей по некоторым уровням координации.
моторной системы. Эти внутренние обратные связи важны не только для стабильного осуществления программы действия, но также для процесса двигательного обучения. Аналогично схеме реафферентации, по Хольсту и Миттельштедту (см. [7]), мы можем предположить, что в каждом из этих уровней двигательной координации образуется и на короткое время складывается как бы копия эфферентной команды движения, которая имеется в распоряжении для сравнения с обратной сигнализацией из нижележащих уровней координации. Рис. 1 демонстрирует принципиальную единую структуру этих внутренних обратных связей. В данном примере от импульсного паттерна подпрограммы 1-го порядка образуется копия, которая сравнивается с обратной информацией о результате этого импульсного паттерна в подпрограммном уровне 2-го порядка и, в случае несогласованности, вызывает изменения импульсного паттерна 1-го порядка. Так, на основе имеющегося состава информации уже на каждом иерархически расположенном уровне координации в центральной нервной системе осуществляется внутренний контроль реализации данного импульсного кода, который обеспечивает соблюдение нужной величины, заданной общей программой. Этот механизм внутренних обратных связей, изображенный на некоторых уровнях координации, имеет следующий вид (рис. 2).
Сегодня нет сомнений, что существуют морфофизиологические основы для этих внутренних обратных связей. Так, две трети восходящих спинноцеребеллярных нервных путей не поставляют специфической информации по модальности и по пространству, а лишь информацию об активности внутри нижележащих двигательных центров и междунейрональных пулов. Оскарссон [8] в 1973 г. указывал на то, что большая часть проходящих между корой, таламусом и мозжечком связей (рис. 3) обеспечивает контроль и корректуру через внутренние обратные связи от коры головного мозга, как верхнего программирующего уровня вытекающих эфферентных импульсов. То же самое относится к ядерным областям нижней оливы, вентрального ретикулярного ядра и другим областям. Копия двигательной команды
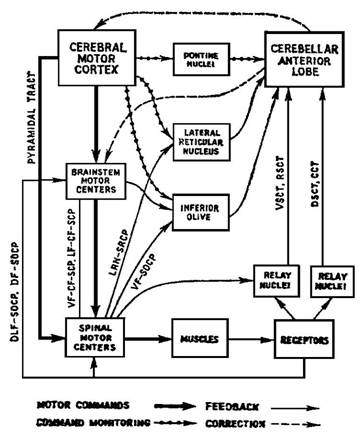
Рис. 3. Некоторые связи между моторной корой головного мозга, передней долей мозжечка и низшими двигательными центрами (Oscarsson, 1973).
118
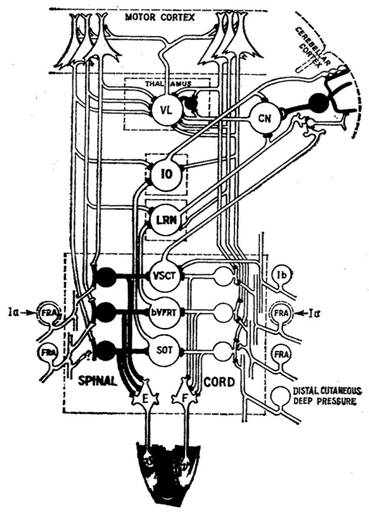
Рис. 4. Схематическое изображение компараторных петель моторной системы по Сцентаготай (1974).
VL — вентролатеральное ядро зрительного бугра; CN — ядро мозжечка; 10 —нижняя олива; LRN — латеральное ретикулярное ядро; VSCT — передний спннномозжечковый путь; VFRT — двухсторонний передний путь флексорного рефлекса; SOT — спиннооливарный путь; FRA — афферентные пути флексорного рефлекса.
или импульсного паттерна на данном координационном уровне имеется в распоряжении не только для сравнения с информациями внешних обратных связей (экстеро- и проприорецепторов), но она является одновременно контрольным паттерном для внутренней об ратной связи ближайшего низшего программирующей уровня, и этот контрольные процесс с обратными связями повторяется на всех низших уровнях моторной системы. Сцентаготай [9] по называет в новом изображении, что эти «компараторные петли» опускаются до уровня спинного мозга (рис. 4).
Исследуя роль кинестетического чувства в управлении произвольными движениями у больных с афферентными парезами, Коц и Найдин [3] делают подобный вывод о существовании афферентации команды. К этому представлении: подходит и то, что у человека было обнаружено исправление ошибок движений, которые, очевидно, не основываются на обратной связи о результате движений. Исследуя следящие движения, Хиггинс и Энжель (1970) описали время исправления ошибок, которое короче времени проприорецептивной реакции. Они заключили, что «испытуемые в состоянии внутренне следить за их собственным поведением, сравнивая актуальные двигательные команды с величиной, которая при данных условиях представляет собой правильную или подходящую команду. Если актуальная и правильная команды не совпадают, то ответная реакция может быть заторможена прежде, чем от периферии вступает обратная связь». Предполагаем, что и другие авторы получили такие же данные.
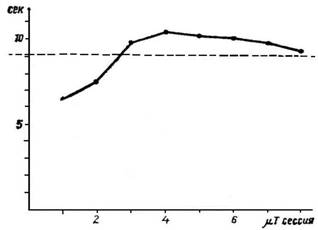
Рис. 5. Изменение продолжительности идеомоторного выполнения сложного навыка во время последовательных опытов.
Такие внутренние обратные связи имеют большое значение для стабильного и приспособленного к ситуации протекания всех навыков. Они являются важным звеном в нейрофизиологическом механизме программированного управления двигательных актов. Но они являются также нейрофизиологической основой эффектов идеомоторной тренировки. При идеомоторном выполнении двигательного навыка, так же как и при действительном движении, осуществляется превращение программы действия в иннервационные паттерны подпрограмм и двигательных элементов (см. рис. 2), как вытекает из токов действия в мышцах, наличие которых во многих случаях доказуемо, и из взаимосвязных изменений вегетативных функций. При этом осуществляются
119
такие внутренние обратные связи на различных уровнях центрального моторного аппарата, которые способствуют более быстрому обучению и стабилизации новой программы действия. «Оживленность» и «близость к реальности воображения идеомоторных движений определяют объем включенных во «внутреннюю реализацию» уровней и таким образом эффективность идеомоторной тренировки. Конечно, этот внутренний механизм корректирования, каким бы он ни был важным для развития и сохранения навыков, поставлен в узкие рамки как самостоятельный фактор. Его можно эффективно использовать только тогда, когда уже существуют подобные навыки и двигательный опыт, или, другими словами, предварительно программированные иннервационные паттерны различных координационных уровней и если временами осуществляется подкрепление внешними реафферентациями от действительно выполненного навыка, т. е. от результата реального движения.
Экспериментальные исследования во время идеомоторной тренировки позволяют нам более глубоко проникнуть в динамические процессы и механизмы отдельных шагов центральной реализации программы действия. Рассмотрим временную структуру идеомоторного тренирующегося навыка. Как вытекает из рис. 5, временная структура идеомоторного воспроизведения навыка отнюдь не должна совпадать со временной структурой его действительного выполнения. Скорее всего она определяется словесной (посторонней или собственной) инструкцией, а также видом и тренировочной стадией воображаемого выполнения навыка. В предложенном примере общее время идеомоторной тренировки навыка закономерно изменяется во время последовательных опытов и постепенно приближается ко времени действительного выполнения. Но, как показывает рис. 6, можно словесными инструкциями обратить внимание тренирующегося на отдельные элементы двигательной структуры, в результате чего удлиняется общее время вследствие временного расширения отдельных элементов. В обоих случаях заданная испытуемому задача заключается в выработке или уточнении нового навыка, т. е. в улучшении или новообразовании сложных программ двигательной координации. Но так же, как показано на рис. 7, я могу идеомоторно упражнять ритм однообразных движений в заданное время. В данном случае стоит задача не выработать новую координацию движений, а повторить определенное число движений в заданное время.
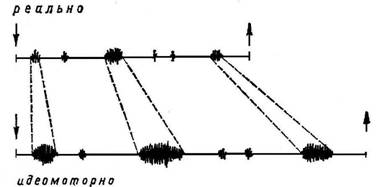
Рис. 6. Удлинение продолжительности идеомоторно воспроизведенного сложного навыка вследствие растяжения отдельных компонентов.
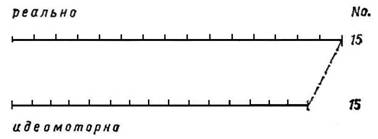
Рис. 7. Укорочение продолжительности идеомоторного воспроизведения тождественных навыков без изменения их числа (No).
Как известно, каждая программа действия, кроме двигательных, включает также вегетативные иннервационные компоненты, которые обеспечивают опережающее снабжение предстоящего моторного акта. Поэтому как показатель степени включения низших координационных уровней иерархической архитектуры моторной системы в ЦНС во время идеомоторной тренировки наряду с появлением мышечных напряжений и электромиографических потенциалов может служить также появление изменений вегетативных функций. Регистрируя их, мы имеем возможность объективно и наглядно уловить временную динамику и частично внутреннюю структуру воображаемого. Вот пример. Испытуемый получил задачу идеомоторно воспроизводить в определенной частоте структуру движений, которой он хорошо владеет, по виду ее выполнения крепко связанной с определенной частотой дыхания (в примере на рис. 8 дело касается плавательных движений). По дыхательной частоте (R) мы можем проследить действительное выполнение этой инструкции, т. е. реализацию в словесной области через некоторые уровни центрального программирования движений осуществляемых процессов, и количественно оценить ее по определенным признакам,— в нашем примере по соблюдению словесно заданной частоты.
Итак, современное состояние наших нейрофизиологических знаний о принципиальной архитектуре в функциональных системах организованных процессов управления и регулирования в моторной системе позволяет нам высказаться о гипотетических нейрофизиологических механизмах, которые лежат в основе доказанных эффектов идеомоторной тренировки. Такая рабочая гипотеза о системе внутренних обратных
120
связей дополнительно открывает концепциональный и экспериментальный подход к специальному внутреннему функциональному принципу высшей нервной деятельности человека. У животных система внутренних обратных связей уже существует в полной мере. Они тоже располагают всеми составными частями «внутреннего» программирования движений и нужными для их реализации механизмами обратной связи и исправления. Они располагают способностью «внутренней» антиципации движений, правда в обширном, с актуальной ситуацией связанном воображаемом пространстве. Можно предположить, что у них внутренние обратные связи являются в основном только конституционными элементами сложной внутренней и внешней общей обратной связи. У человека во время эволюции создалась другая ситуация. У него внутренние обратные связи по лучили самостоятельное значение: они стали основой развития словесных функций, которые формировались вследствие рабочей деятельности и деятельной общественной коммуникации с другими людьми. Тем самым на уровне человека такая система внутренних механизмов обратной связи приобрела новое качество. Образование отвлеченных представлений «при заторможенном двигательном конечном звене», на которое И. М. Сеченов указывал еще в 1863 г., создало возможность отражения не только актуальных, но и бывших и будущих событий, действий и установок в воображаемом пространственно-временном континууме. Сознательное проигрывание идеомоторных актов представляет собой частный случай подобных процессов, в ходе которых опробывается и упражняется эфферентный путь программы действия, т. е. исполнительная часть поведения.
Так, идеомоторная тренировка по отношению к ее конкретной структуре стоит между действительным выполнением навыка и его словесным проектом. Это нам открывает возможность, исследуя конкретные нейрофизиологические процессы во время идеомоторной тренировки, глубже проникнуть в програничную область архитектуры и динамики сложных психофизиологических процессов управления и регулирования, которые лежат в основе реализации актов поведения человека.

Рис. 8. Ускорение дыхательной частоты во время идеомоторного воспроизведения плавания на определенную дистанцию в заданное время.
R — время (сек) отдельных вдохов.
1. Анохин П. К. Биология и нейрофизиология условного рефлекса. — М., 1968.
2. Бернштейн Н. А. Очерки по физиологии движений и физиологии активности. — М., 1966.
3. Коц Я. М. и Найдин В. Л. О роли кинестетического чувства в управлении непроизвольными движениями. — Вопросы психологии, 1966, № 5, с. 114—122
4. Павлов И. П. Поли. собр. соч., 3, кн. 2. — М.; Л., 1951.
5. Сеченов И. М. Рефлексы головного мозга. — СПб., 1863.
6. Haggins J. R. and Angel R. W. Correction of tracking errors without sensory feedback. — J. exp. Psychol., 1970, 84, p. 412—426.
7. Hoist E. and v. Mittelstaedt H. Das Reafferenzorinzip Naturwissenschaiten, 1950, 37, p. 464—476.
8. Oscarsson O. Functional organisation of spinocerebellar paths. In: Iggo A. (Ed.). Handbook of Sensory Physiology, vol. 11. Somatosensory System, S. 339—380. — Berlin: Springer-Verlag, 1973.
9. Szentagothai J. and Arbib At. A. Conceptual models of neural organization. Neurosci. — Res. Prograrnm Bull., 1974, 12.